

Phospholemman, a single-span membrane protein, is an accessory protein of Na,K-ATPase in cerebellum and choroid plexus
- PMID: 12657675
- PMCID: PMC6742001
- DOI: 10.1523/JNEUROSCI.23-06-02161.2003
Phospholemman, a single-span membrane protein, is an accessory protein of Na,K-ATPase in cerebellum and choroid plexus
Abstract
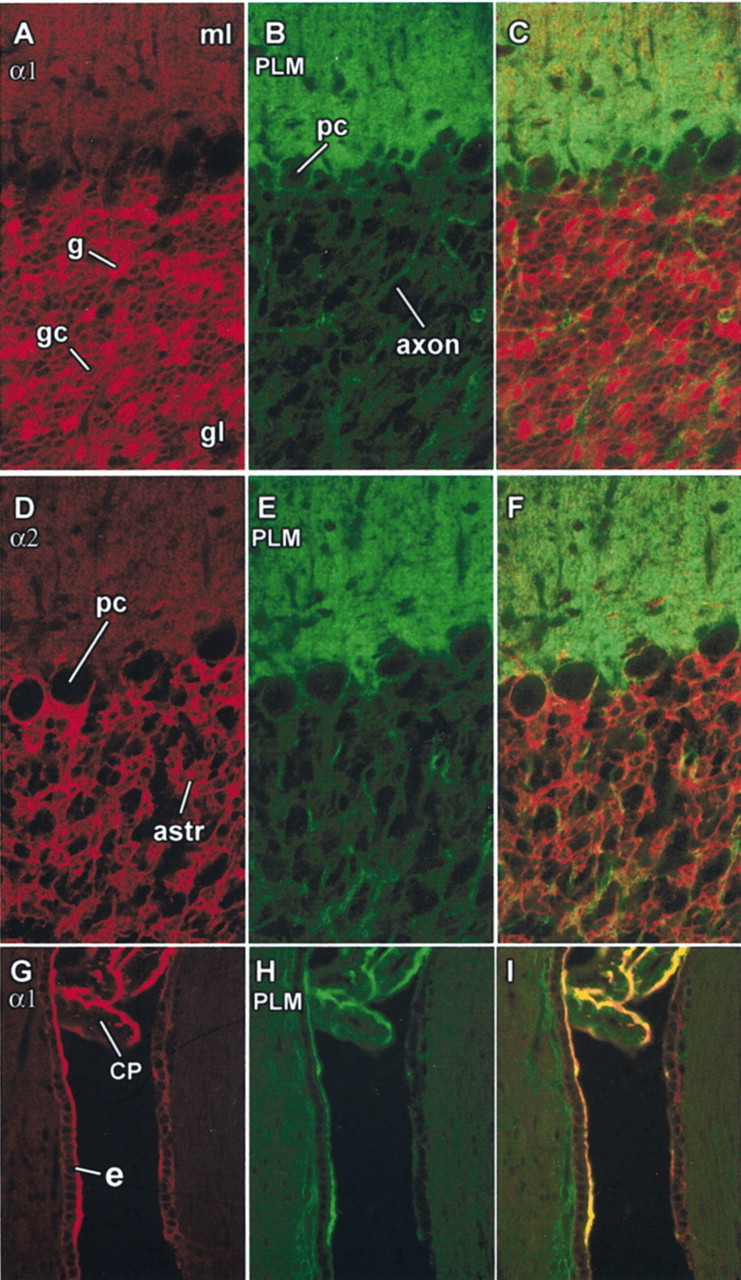
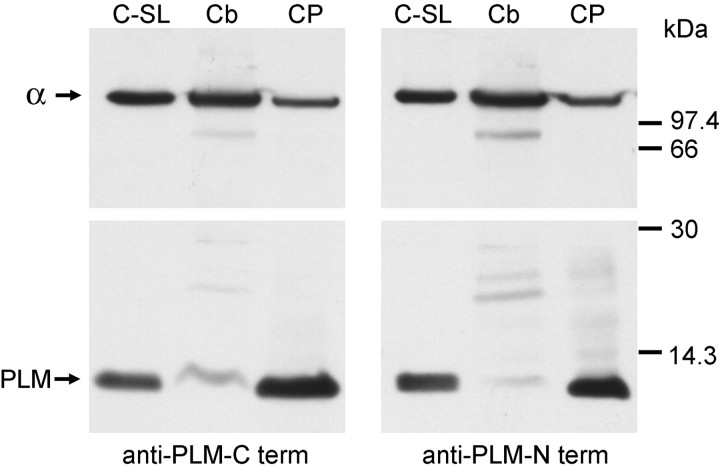
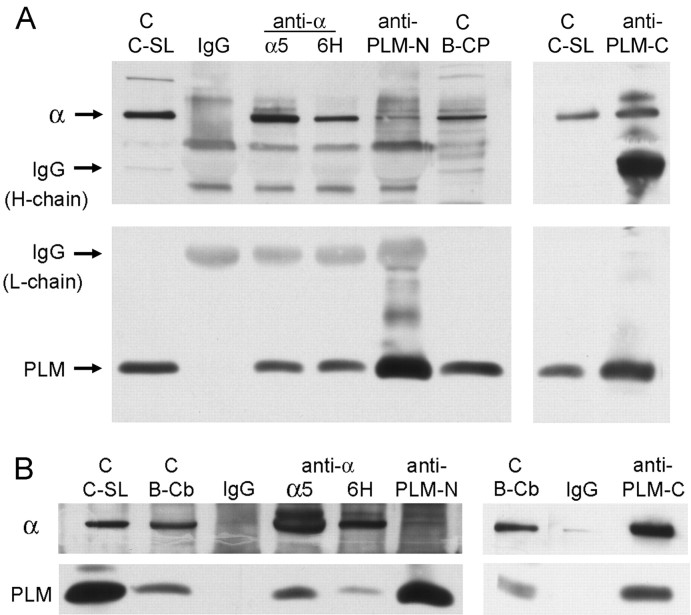
Phospholemman (FXYD1) is a homolog of the Na,K-ATPase gamma subunit (FXYD2), a small accessory protein that modulates ATPase activity. Here we show that phospholemman is highly expressed in selected structures in the CNS. It is most abundant in cerebellum, where it was detected in the molecular layer, in Purkinje neurons, and in axons traversing the granule cell layer. Phospholemman was particularly enriched in choroid plexus, the organ that secretes CSF in the ventricles, where it colocalized with Na,K-ATPase in the apical membrane. It was also enriched, with Na,K-ATPase, in certain tanycytes or ependymal cells of the ventricle wall. Two different experimental approaches demonstrated that phospholemman physically associated with the Na,K-ATPase in cerebellum and choroid plexus: the proteins copurified after detergent treatment and co-immunoprecipitated from solubilized crude membranes using either anti-phospholemman or anti-Na,K-ATPase antibodies. Phospholemman antibodies precipitated all three Na,K-ATPase alpha subunit isoforms (alpha1-alpha3) from cerebellum, indicating that the interaction is not specific to a particular alpha isoform and consistent with the presence of phospholemman in both neurons and glia. Antibodies against the C-terminal domain of phospholemman reduced Na,K-ATPase activity in vitro without effect on Na+ affinity. At least two other FXYD family members have been detected in the CNS, suggesting that additional complexity of sodium pump regulation will be found.
Figures





Similar articles
-
FXYD proteins stabilize Na,K-ATPase: amplification of specific phosphatidylserine-protein interactions.J Biol Chem. 2011 Mar 18;286(11):9699-712. doi: 10.1074/jbc.M110.184234. Epub 2011 Jan 12. J Biol Chem. 2011. PMID: 21228272 Free PMC article.
-
Identification of a phospholemman-like protein from shark rectal glands. Evidence for indirect regulation of Na,K-ATPase by protein kinase c via a novel member of the FXYDY family.J Biol Chem. 2000 Nov 17;275(46):35969-77. doi: 10.1074/jbc.M005168200. J Biol Chem. 2000. PMID: 10961995
-
Phosphorylation of phospholemman (FXYD1) by protein kinases A and C modulates distinct Na,K-ATPase isozymes.J Biol Chem. 2008 Jan 4;283(1):476-486. doi: 10.1074/jbc.M705830200. Epub 2007 Nov 8. J Biol Chem. 2008. PMID: 17991751
-
[FXYD proteins: novel regulators of Na,K-ATPase].Med Sci (Paris). 2006 Jun-Jul;22(6-7):633-8. doi: 10.1051/medsci/20062267633. Med Sci (Paris). 2006. PMID: 16828040 Review. French.
-
FXYD proteins: new regulators of Na-K-ATPase.Am J Physiol Renal Physiol. 2006 Feb;290(2):F241-50. doi: 10.1152/ajprenal.00126.2005. Am J Physiol Renal Physiol. 2006. PMID: 16403837 Review.
Cited by
-
Endogenous Na+, K+-ATPase inhibitors and CSF [Na+] contribute to migraine formation.PLoS One. 2019 Jun 7;14(6):e0218041. doi: 10.1371/journal.pone.0218041. eCollection 2019. PLoS One. 2019. PMID: 31173612 Free PMC article.
-
Multiple Na,K-ATPase Subunits Colocalize in the Brush Border of Mouse Choroid Plexus Epithelial Cells.Int J Mol Sci. 2021 Feb 4;22(4):1569. doi: 10.3390/ijms22041569. Int J Mol Sci. 2021. PMID: 33557294 Free PMC article.
-
Basis of chloride transport in ciliary epithelium.J Membr Biol. 2004 Jul 1;200(1):1-13. doi: 10.1007/s00232-004-0688-5. J Membr Biol. 2004. PMID: 15386155 Review.
-
FXYD proteins and sodium pump regulatory mechanisms.J Gen Physiol. 2021 Apr 5;153(4):e202012633. doi: 10.1085/jgp.202012633. J Gen Physiol. 2021. PMID: 33688925 Free PMC article. Review.
-
Brain region-specific expression of Fxyd1, an Mecp2 target gene, is regulated by epigenetic mechanisms.J Neurosci Res. 2011 Jun;89(6):840-51. doi: 10.1002/jnr.22608. Epub 2011 Mar 10. J Neurosci Res. 2011. PMID: 21394759 Free PMC article.
References
-
- Arystarkhova E, Sweadner KJ. Isoform-specific monoclonal antibodies to Na-K-ATPase α subunits: evidence for a tissue-specific post-translational modification of the α subunit. J Biol Chem. 1996;271:23407–23417. - PubMed
-
- Arystarkhova E, Wetzel RK, Asinovski NK, Sweadner KJ. The γ subunit modulates Na+ and K+ affinity of the renal Na,K-ATPase. J Biol Chem. 1999;274:33183–33185. - PubMed
-
- Arystarkhova E, Donnet C, Asinovski NK, Sweadner KJ. Differential regulation of renal Na,K-ATPase by splice variants of the γ subunit. J Biol Chem. 2002;277:10162–10172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases