

Adenylate cyclase 1 as a key actor in the refinement of retinal projection maps
- PMID: 12657682
- PMCID: PMC6742000
- DOI: 10.1523/JNEUROSCI.23-06-02228.2003
Adenylate cyclase 1 as a key actor in the refinement of retinal projection maps
Abstract
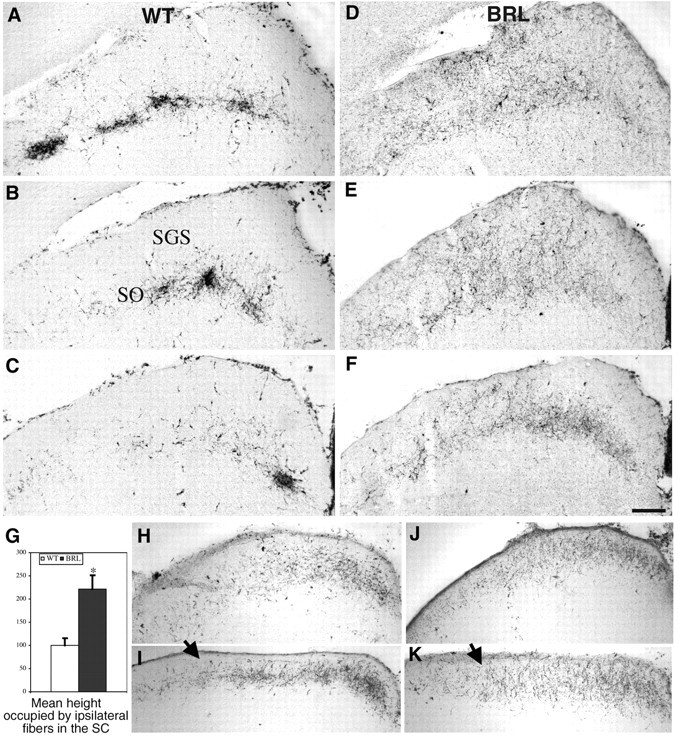
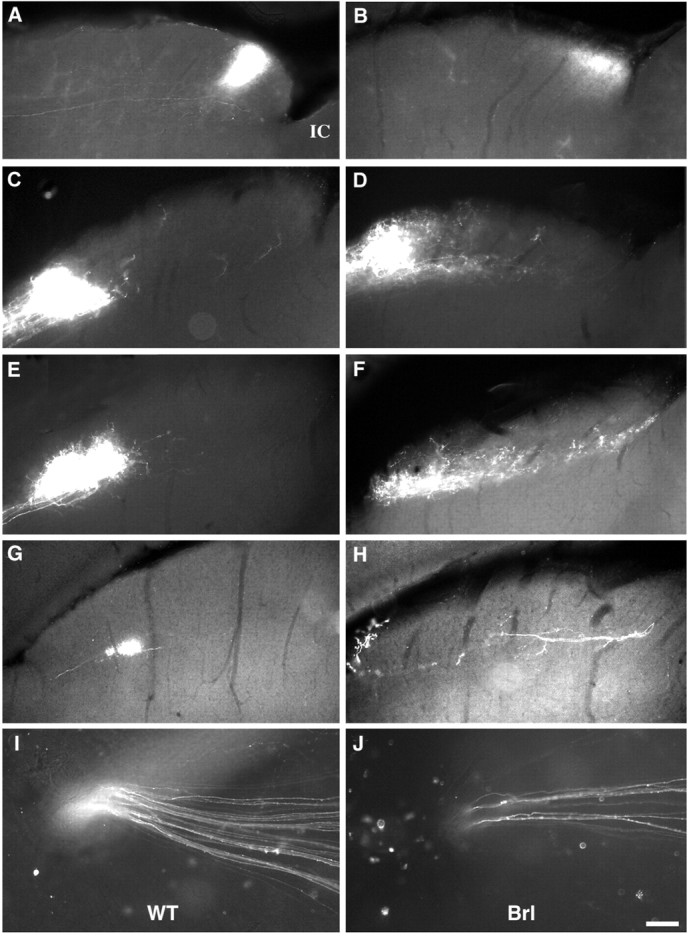
cAMP occupies a strategic position to control neuronal responses to a large variety of developmental cues. We have analyzed the role of calcium-stimulated adenylate cyclase 1 (AC1) in the development of retinal topographic maps. AC1 is expressed in retinal ganglion cells (RGCs) from embryonic day 15 to adulthood with a peak during the first postnatal week. At that time, the other calcium-stimulated AC, AC8, is expressed in the superior colliculus (SC) but not in the RGCs. In mice of the barrelless strain, which carry an inactivating mutation of the AC1 gene, calcium-stimulated AC activity is reduced by 40-60% in the SC and retina. RGC projection maps were analyzed with a variety of anterograde and retrograde tracers. After an initially normal development until postnatal day 3, retinal fibers from the ipsilateral and contralateral eye fail to segregate into eye-specific domains in the lateral geniculate nucleus and the SC. Topographic defects in the fine tuning of the retinotectal and retinogeniculate maps are also observed with abnormalities in the confinement of the retinal axon arbors in the anteroposterior and mediolateral dimensions. This is attributable to the lack of elimination of misplaced axon collaterals and to the maintenance of a transient ipsilateral projection. These results establish an essential role of AC1 in the fine patterning of the retinal map. Calcium-modulated cAMP production in the RGCs could constitute an important link between activity-dependent changes and the anatomical restructuring of the retinal terminal arbors within central targets.
Figures






References
-
- Abdel-Majid RM, Leong WL, Schalkwyk LC, Smallman DS, Wong ST, Storm DR, Fine A, Dobson MJ, Guernsey DL, Neumann PE. Loss of adenylate cyclase I activity disrupts patterning of mouse somatosensory cortex. Nat Genet. 1998;19:289–291. - PubMed
-
- Brooks SP, Storey KB. Bound and determined: a computer program for making buffers of defined ion concentrations. Anal Biochem. 1992;201:119–126. - PubMed
-
- Cali JJ, Zwaagstra JC, Mons N, Cooper DM, Krupinski J. Type VIII adenylate cyclase: a Ca2+/calmodulin-stimulated enzyme expressed in discrete regions of rat brain. J Biol Chem. 1994;269:12190–12195. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases