

Rap1p telomere association is not required for mitotic stability of a C(3)TA(2) telomere in yeast
- PMID: 12660174
- PMCID: PMC152898
- DOI: 10.1093/emboj/cdg154
Rap1p telomere association is not required for mitotic stability of a C(3)TA(2) telomere in yeast
Abstract
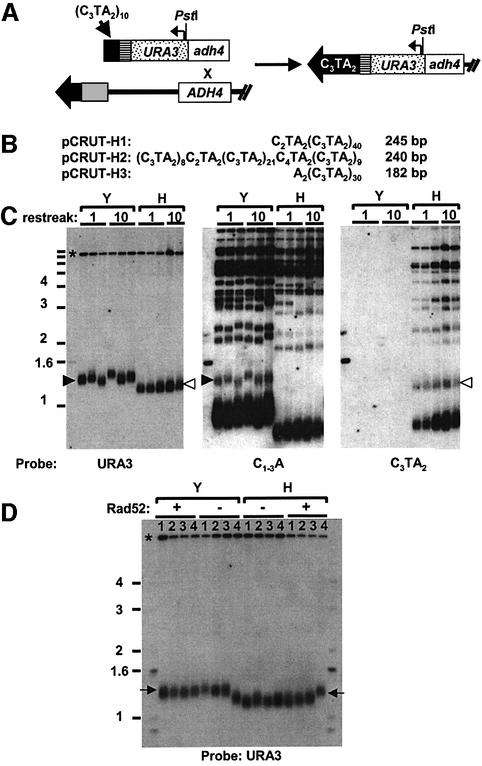
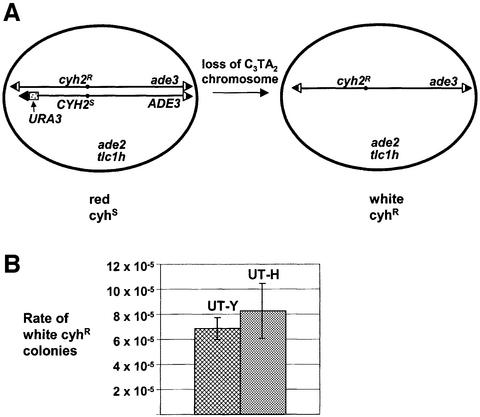
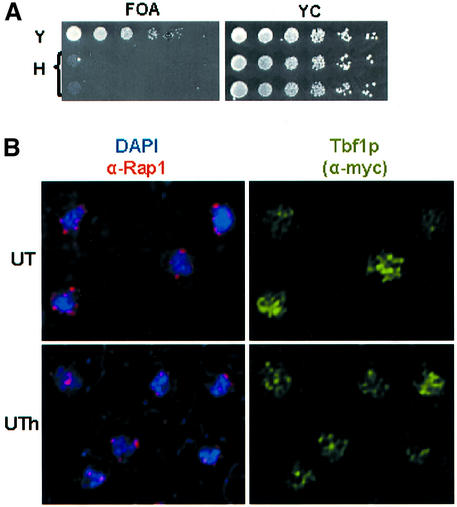
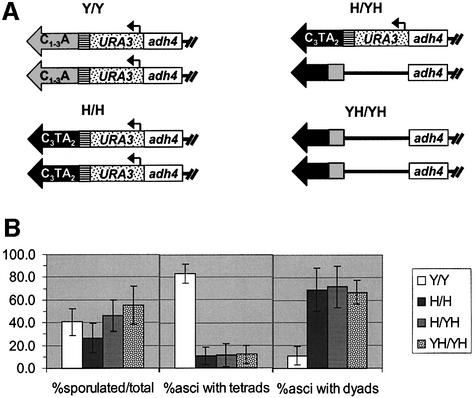
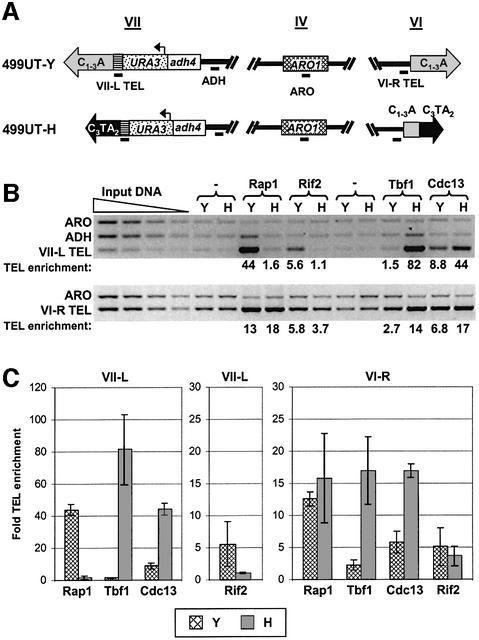
Telomeric DNA usually consists of a repetitive sequence: C(1-3)A/TG(1-3) in yeast, and C(3)TA(2)/T(2)AG(3) in vertebrates. In yeast, the sequence-specific DNA- binding protein Rap1p is thought to be essential for telomere function. In a tlc1h mutant, the templating region of the telomerase RNA gene is altered so that telomerase adds the vertebrate telomere sequence instead of the yeast sequence to the chromosome end. A tlc1h strain has short but stable telomeres and no growth defect. We show here that Rap1p and the Rap1p-associated Rif2p did not bind to a telomere that contains purely vertebrate repeats, while the TG(1-3) single-stranded DNA binding protein Cdc13p and the normally non-telomeric protein Tbf1p did bind this telomere. A chromosome with one entirely vertebrate-sequence telomere had a wild-type loss rate, and the telomere was maintained at a short but stable length. However, this telomere was unable to silence a telomere-adjacent URA3 gene, and the strain carrying this telomere had a severe defect in meiosis. We conclude that Rap1p localization to a C(3)TA(2) telomere is not required for its essential mitotic functions.
Figures
References
-
- Aparicio O.M., Billington,B.L. and Gottschling,D.E. (1991) Modifiers of position effect are shared between telomeric and silent mating-type loci in S.cerevisiae. Cell, 66, 1279–1287. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
