

The number of vertebrate repeats can be regulated at yeast telomeres by Rap1-independent mechanisms
- PMID: 12660175
- PMCID: PMC152899
- DOI: 10.1093/emboj/cdg155
The number of vertebrate repeats can be regulated at yeast telomeres by Rap1-independent mechanisms
Abstract
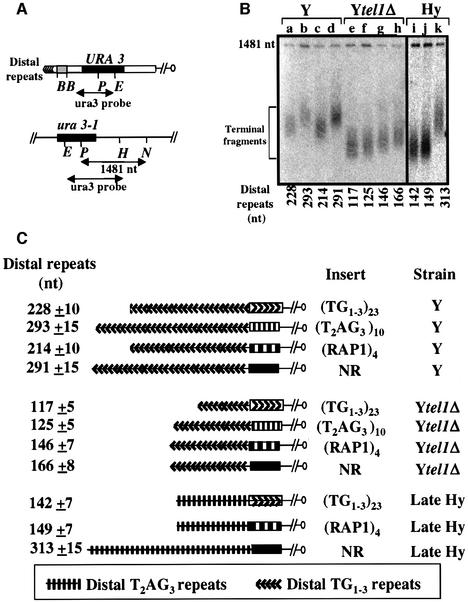
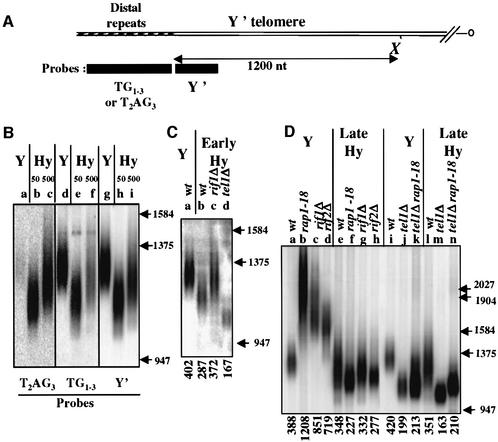
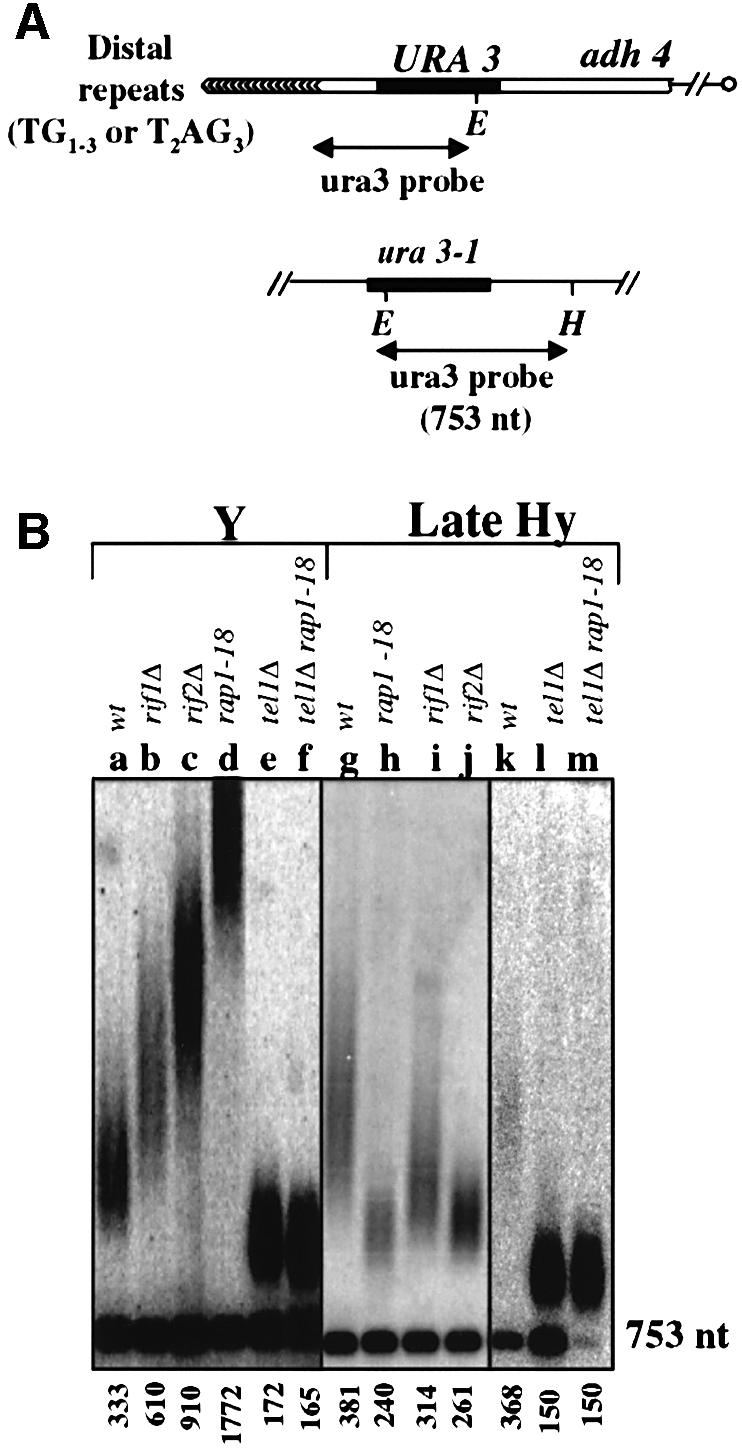
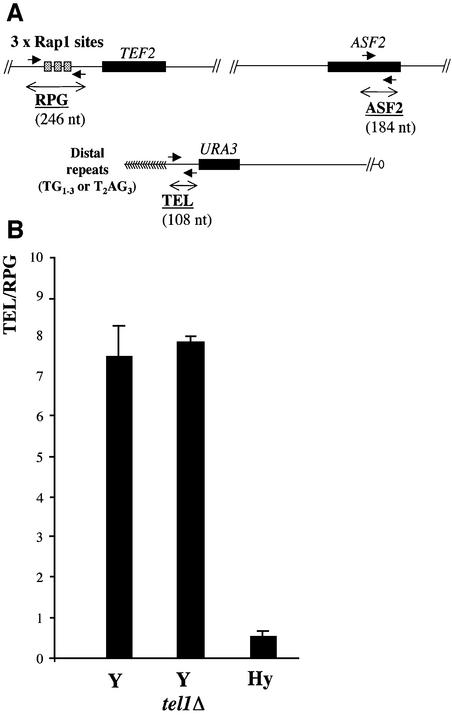
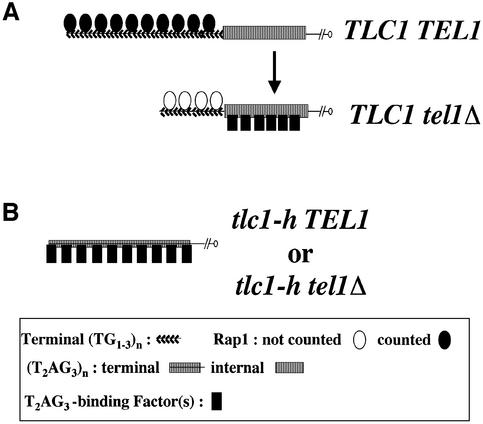
The number of telomeric DNA repeats at chromosome ends is maintained around a mean value by a dynamic balance between elongation and shortening. In particular, proteins binding along the duplex part of telomeric DNA set the number of repeats by progressively limiting telomere growth. The paradigm of this counting mechanism is the Rap1 protein in Saccharomyces cerevisiae. We demonstrate here that a Rap1-independent mechanism regulates the number of yeast telomeric repeats (TG(1-3)) and of vertebrate repeats (T(2)AG(3)) when TEL1, a yeast ortholog of the human gene encoding the ATM kinase, is inactivated. In addition, we show that a T(2)AG(3)-only telomere can be formed and maintained in humanized yeast cells carrying a template mutation of the gene encoding the telomerase RNA, which leads to the synthesis of vertebrate instead of yeast repeats. Genetic and biochemical evidences indicate that this telomere is regulated in a Rap1-independent manner, both in TEL1 and in tel1Delta humanized yeast cells. Altogether, these findings shed light on multiple repeat-counting mechanisms, which may share critical features between lower and higher eukaryotes.
Figures
References
-
- Ancelin K., Brunori,M., Bauwens,S., Koering,C.E., Brun,C., Ricoul,M., Pommier,J.P., Sabatier,L. and Gilson,E. (2002) Targeting assay to study the cis functions of human telomeric proteins: evidence for inhibition of telomerase by TRF1 and for activation of telomere degradation by TRF2. Mol. Cell. Biol., 22, 3474–3487. - PMC - PubMed
-
- Bianchi A. and de Lange,T. (1999) Ku binds telomeric DNA in vitro. J. Biol. Chem., 274, 21223–21227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
