

Structural organization of authentic, mature HIV-1 virions and cores
- PMID: 12660176
- PMCID: PMC152888
- DOI: 10.1093/emboj/cdg143
Structural organization of authentic, mature HIV-1 virions and cores
Abstract
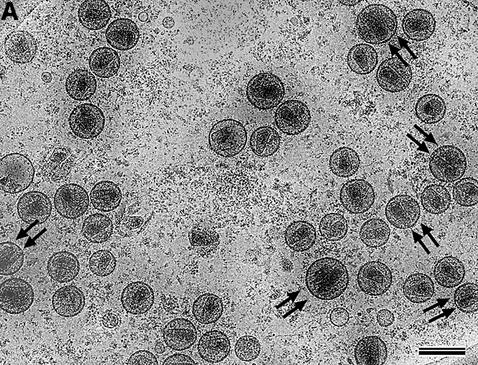
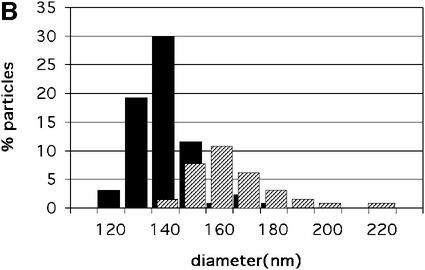
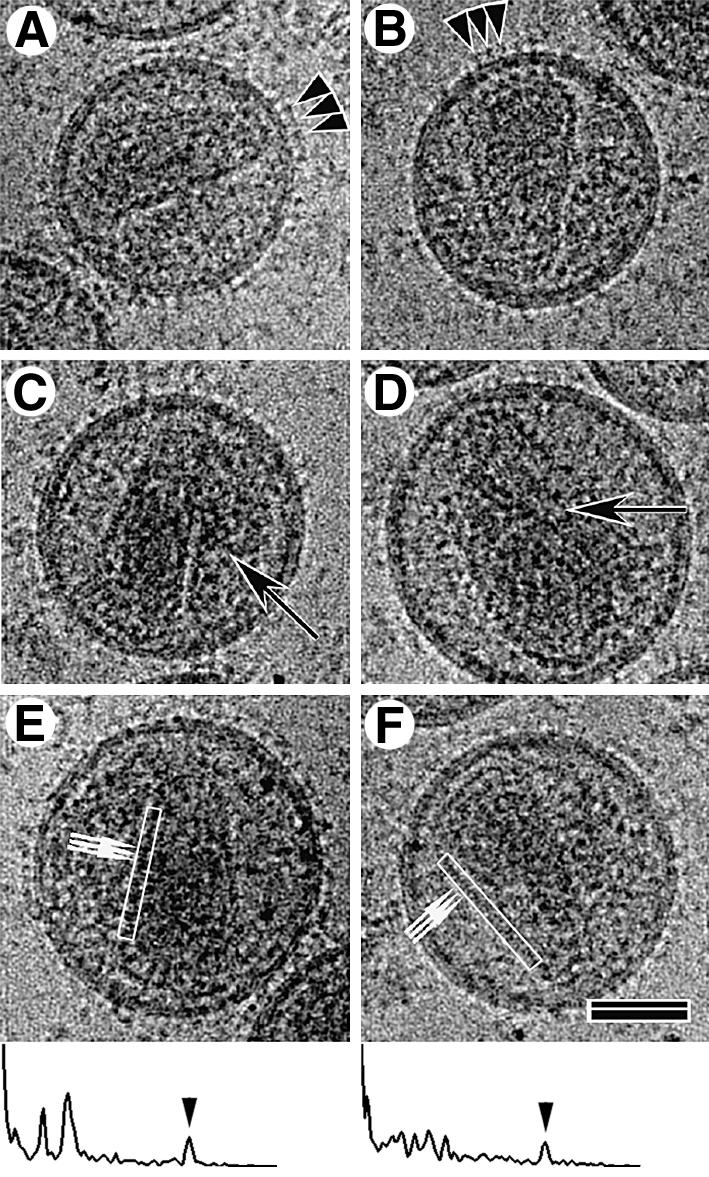
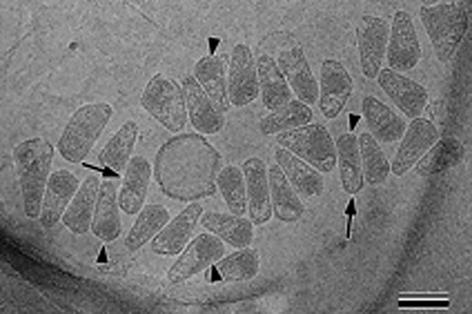
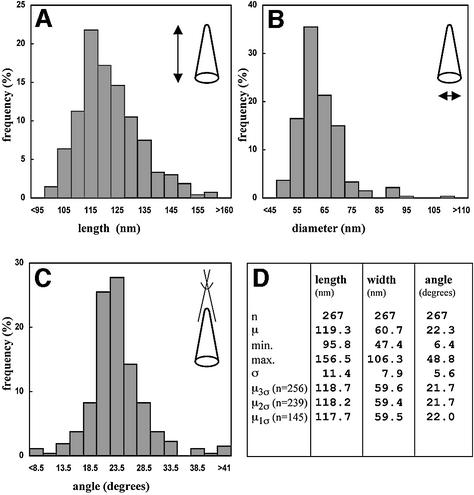
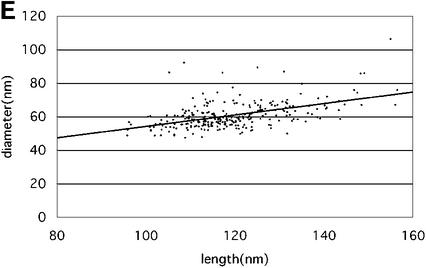
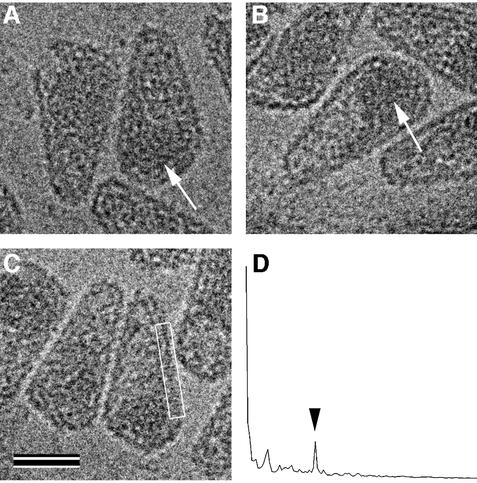
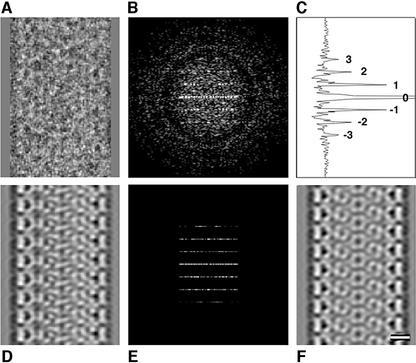
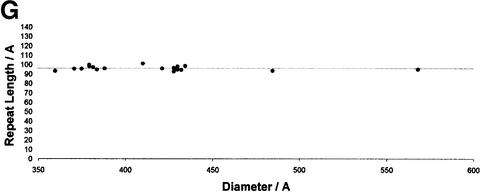
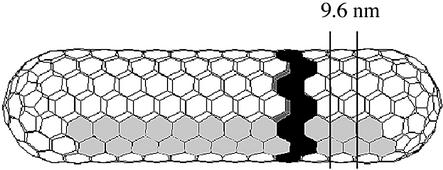
Mature, infectious HIV-1 particles contain a characteristic cone-shaped core that encases the viral RNA and replication proteins. The architectures of mature virions and isolated cores were studied using cryo-electron microscopy. The average size ( approximately 145 nm) of the virion was unchanged during maturation. Most virions contained a single core but roughly one-third contained two or more cores. Consideration of the capsid protein concentration during core assembly indicated that core formation in vivo is template-mediated rather than concentration-driven. Although most cores were conical, 7% were tubular. These displayed a stacked-disc arrangement with 7-, 8-, 9- or 10-fold axial symmetry. Layer line filtration of these images showed that the capsid subunit arrangement is consistent with a 9.6 nm hexamer resembling that previously seen in the helical tubes assembled from purified capsid protein. A common reflection (1/3.2 nm) shared between the tubular and conical cores suggested they share a similar organization. The extraordinary flexibility observed in the assembly of the mature core appears to be well suited to accommodating variation and hence there may be no single structure for the infectious virion.
Figures
References
-
- Barklis E., McDermott,J., Wilkens,S., Fuller,S. and Thompson,D. (1998) Organization of HIV-1 capsid proteins on a lipid monolayer. J. Biol. Chem., 273, 7177–7180. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
