

Silencing of the baculovirus Op-iap3 gene by RNA interference reveals that it is required for prevention of apoptosis during Orgyia pseudotsugata M nucleopolyhedrovirus infection of Ld652Y cells
- PMID: 12663755
- PMCID: PMC152165
- DOI: 10.1128/jvi.77.8.4481-4488.2003
Silencing of the baculovirus Op-iap3 gene by RNA interference reveals that it is required for prevention of apoptosis during Orgyia pseudotsugata M nucleopolyhedrovirus infection of Ld652Y cells
Abstract
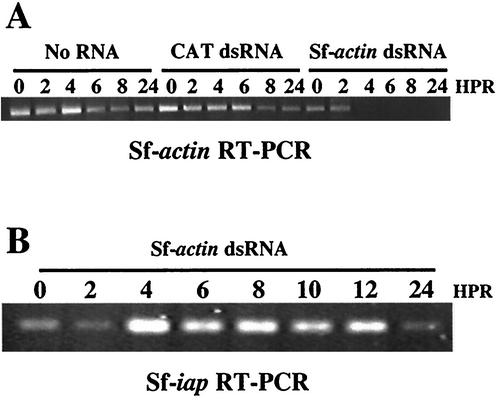
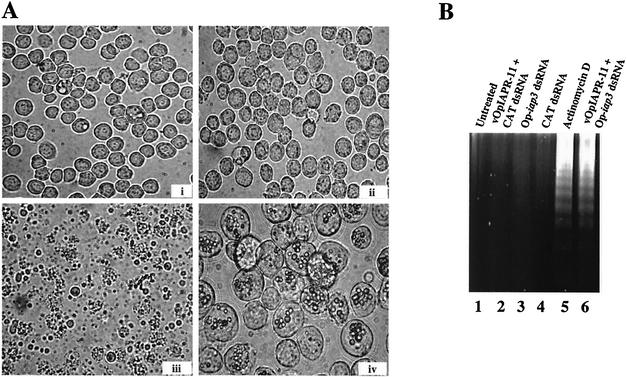
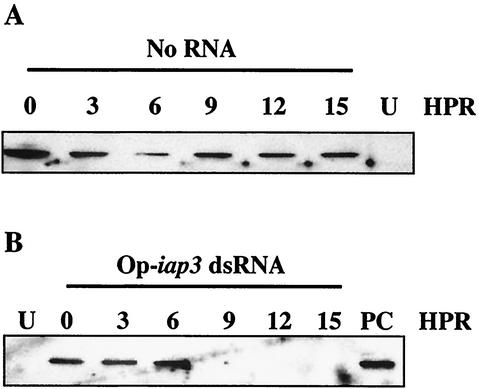
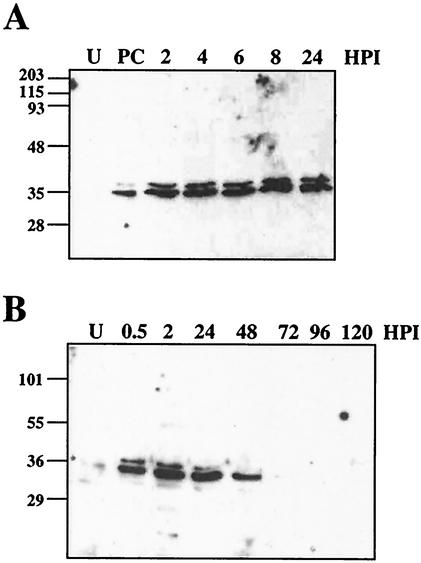
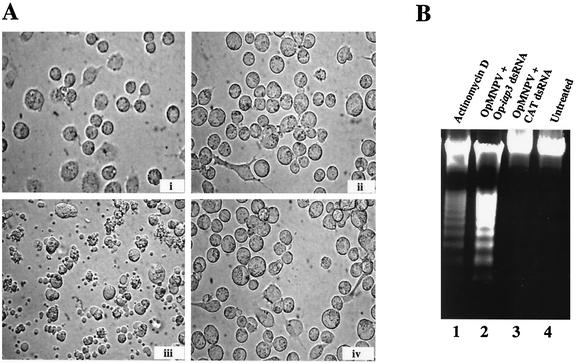
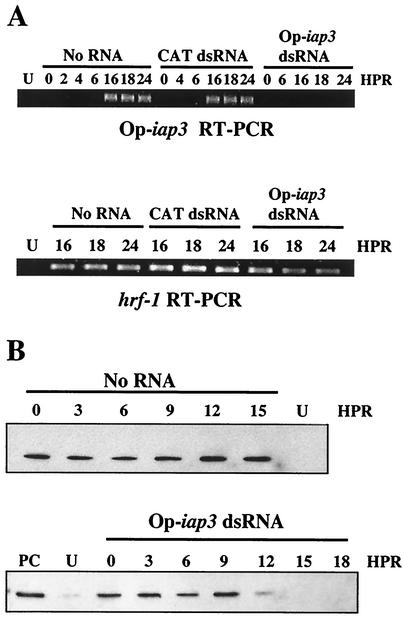
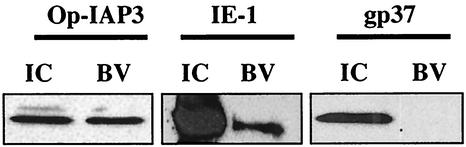
The Op-iap3 gene from the baculovirus Orgyia pseudotsugata M nucleopolyhedrovirus (OpMNPV) inhibits apoptosis induced by a mutant of Autographa californica MNPV (AcMNPV) that lacks the antiapoptotic gene p35, as well as apoptosis induced by a wide range of other stimuli in both mammalian and insect cells. However, the role of Op-iap3 during OpMNPV infection has not been previously examined. To determine the function of the Op-IAP3 protein during OpMNPV infection, we used RNA interference (RNAi) to silence Op-iap3 expression during OpMNPV infection of Ld652Y cells. Infected cells treated with Op-iap3 double-stranded RNA (dsRNA) did not accumulate detectable Op-iap3 mRNA, confirming that the Op-iap3 gene was effectively silenced. Op-IAP3 protein was found to be a component of the budded virion; however, in OpMNPV-infected cells treated with Op-iap3 dsRNA, the Op-IAP3 protein that was introduced by the inoculum virus decreased to almost undetectable levels by 12 h after dsRNA addition. Apoptosis was observed in infected cells treated with Op-iap3 dsRNA beginning at 12 h, and by 48 h, almost all of the cells had undergone apoptosis. These results show for the first time that Op-IAP3 is necessary to prevent apoptosis during OpMNPV infection. In addition, our results demonstrate that the RNAi technique can be an effective tool for studying baculovirus gene function.
Figures
Similar articles
-
Ubiquitin protein ligase activity of the anti-apoptotic baculovirus protein Op-IAP3.Virus Res. 2004 Sep 15;105(1):89-96. doi: 10.1016/j.virusres.2004.04.017. Virus Res. 2004. PMID: 15325084
-
Baculovirus Lymantria dispar multiple nucleopolyhedrovirus IAP2 and IAP3 do not suppress apoptosis, but trigger apoptosis of insect cells in a transient expression assay.Virus Genes. 2012 Oct;45(2):370-9. doi: 10.1007/s11262-012-0783-0. Epub 2012 Jul 15. Virus Genes. 2012. PMID: 22798056
-
Functional analysis of two inhibitor of apoptosis (iap) orthologs from Helicoverpa armigera nucleopolyhedrovirus.Virus Res. 2012 Apr;165(1):107-11. doi: 10.1016/j.virusres.2012.01.012. Epub 2012 Feb 4. Virus Res. 2012. PMID: 22326337
-
Baculoviruses and apoptosis: a diversity of genes and responses.Curr Drug Targets. 2007 Oct;8(10):1069-74. doi: 10.2174/138945007782151405. Curr Drug Targets. 2007. PMID: 17979666 Review.
-
Insect defenses against virus infection: the role of apoptosis.Int Rev Immunol. 2003 Sep-Dec;22(5-6):401-24. doi: 10.1080/08830180305215. Int Rev Immunol. 2003. PMID: 12959752 Review.
Cited by
-
Novel apoptosis suppressor Apsup from the baculovirus Lymantria dispar multiple nucleopolyhedrovirus precludes apoptosis by preventing proteolytic processing of initiator caspase Dronc.J Virol. 2013 Dec;87(23):12925-34. doi: 10.1128/JVI.02065-13. Epub 2013 Sep 25. J Virol. 2013. PMID: 24067961 Free PMC article.
-
Genomic sequencing of Troides aeacus nucleopolyhedrovirus (TraeNPV) from golden birdwing larvae (Troides aeacus formosanus) to reveal defective Autographa californica NPV genomic features.BMC Genomics. 2019 May 27;20(1):419. doi: 10.1186/s12864-019-5713-2. BMC Genomics. 2019. PMID: 31133070 Free PMC article.
-
Expression of baculovirus late and very late genes depends on LEF-4, a component of the viral RNA polymerase whose guanyltransferase function is essential.J Virol. 2006 Apr;80(8):4168-73. doi: 10.1128/JVI.80.8.4168-4173.2006. J Virol. 2006. PMID: 16571832 Free PMC article.
-
Identification and functional analysis of the putative anti-apoptotic gene iap4 of Spodoptera litura nucleopolyhedrovirus.Virus Genes. 2010 Oct;41(2):298-304. doi: 10.1007/s11262-010-0513-4. Epub 2010 Jul 13. Virus Genes. 2010. PMID: 20625923
-
Enhanced recombinant protein production and differential expression of molecular chaperones in sf-caspase-1-repressed stable cells after baculovirus infection.BMC Biotechnol. 2012 Nov 7;12:83. doi: 10.1186/1472-6750-12-83. BMC Biotechnol. 2012. PMID: 23134743 Free PMC article.
References
-
- Ahrens, C. H., R. L. Q. Russell, C. J. Funk, J. T. Evans, S. H. Harwood, and G. F. Rohrmann. 1997. The sequence of the Orgyia pseudotsugata multinucleocapsid nuclear polyhedrosis virus genome. Virology 229:381-399. - PubMed
-
- Bawden, A. L., K. J. Glassberg, J. Diggans, R. Shaw, W. Farmerie, and R. W. Moyer. 2000. Complete genomic sequence of the Amsacta moorei entomopoxvirus: analysis and comparison with other poxviruses. Virology 274:120-139. - PubMed
-
- Bernstein, E., A. A. Caudy, S. M. Hammond, and G. J. Hannon. 2001. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409:363-366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
