

Strain variations in single amino acids of the 86-kilodalton human cytomegalovirus major immediate-early protein (IE2) affect its functional and biochemical properties: implications of dynamic protein conformation
- PMID: 12663783
- PMCID: PMC152111
- DOI: 10.1128/jvi.77.8.4760-4772.2003
Strain variations in single amino acids of the 86-kilodalton human cytomegalovirus major immediate-early protein (IE2) affect its functional and biochemical properties: implications of dynamic protein conformation
Abstract
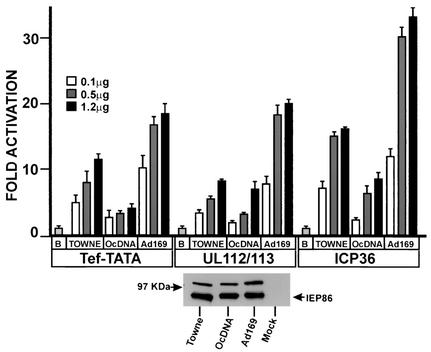
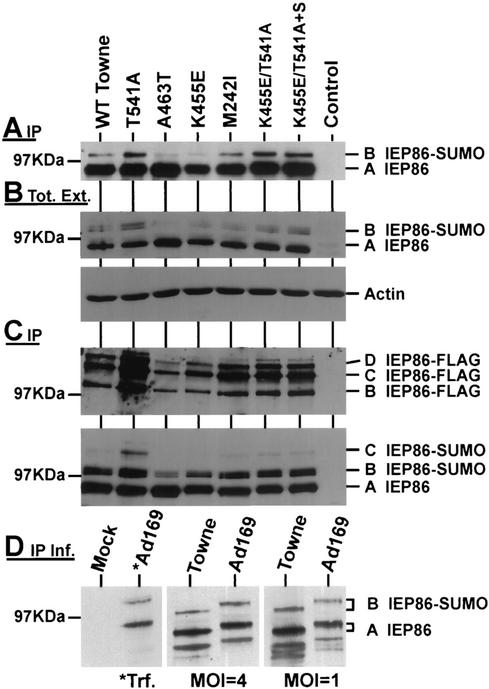
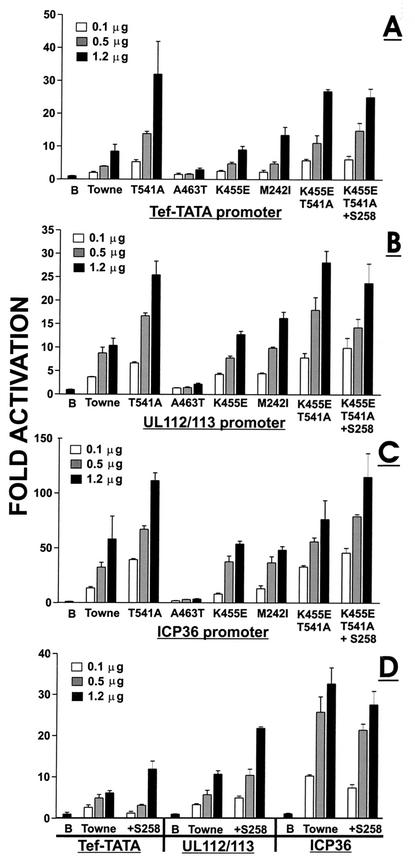
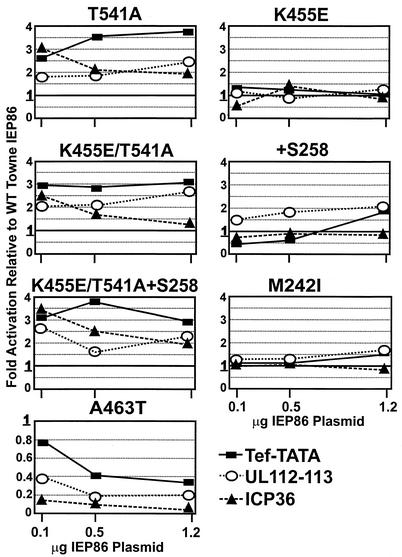
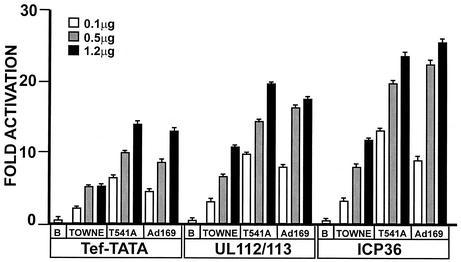
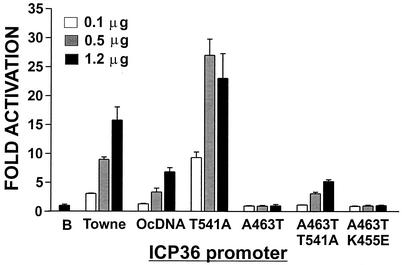
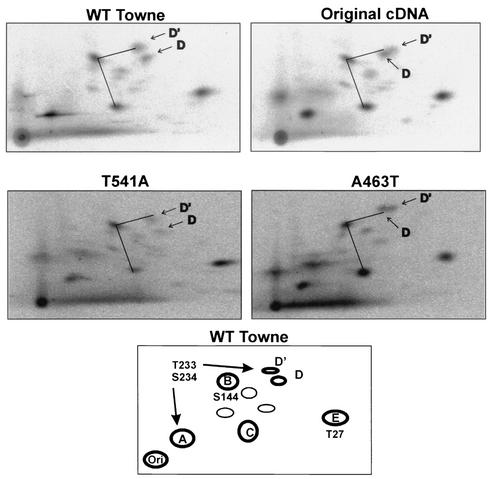
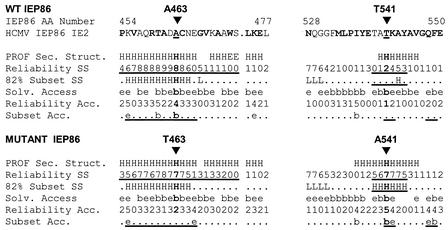
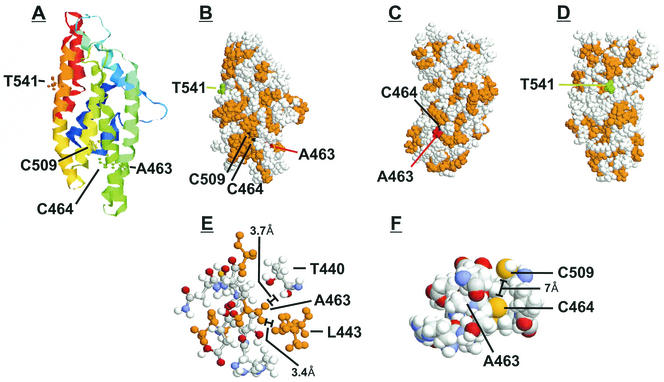
The 86-kDa major immediate-early protein, IEP86 (IE2, IE2(579aa), or ppUL122a), from the Towne and AD169 strains of human cytomegalovirus show four amino acid variations, namely, R68Q, K455E, T541A, and seven consecutive serines beginning at position 258 in Towne and eight serines in AD169. A commonly utilized IEP86 cDNA expression clone (herein called the original cDNA) (E. Baracchini, E. Glezer, K. Fish, R. M. Stenberg, J. A. Nelson, and P. Ghazal, Virology 188:518-529, 1992) shows the Towne R68 and seven serines but contains the AD169 E455 and A541 plus two amino acid mutations, M242I and A463T. In transcriptional activation analyses using several promoters, the IEP86 produced by the original cDNA was 40 to 60% less active than wild-type (WT) Towne IEP86, whereas AD169 IEP86 was two to three times more active than WT Towne IEP86. To determine which amino acid variations or mutations accounted for the differences in transcriptional activation, they were individually tested in the WT Towne IEP86 background. K455E, M242I, and the eighth serine had little effect on transcriptional activation or sumoylation when inserted into the Towne background. T541A significantly increased transcriptional activation on all promoters tested and showed increased sumoylation; T541A is the primary reason that WT AD169 IEP86 has increased activity over WT Towne IEP86. The increased sumoylation seen with T541A was quantitatively reduced to WT Towne levels when the K455E alteration was present, suggesting that K455 may be a sumoylation site or that E455 may cause alterations in the IEP86 structure which affect overall sumoylation. A463T was very deleterious to transcriptional activation and caused reduced sumoylation. The A436T mutation in the original cDNA is partially compensated by the presence of the T541A variation. Phosphopeptide mapping suggests that a threonine at 463 or 541 does not introduce a phosphorylation site. However, the A463T mutation does affect phosphorylation at a distant site, suggesting that it alters the conformation of the protein. Promoter-specific effects were noted with some of the amino acid variations, particularly T541A. Structural modeling is presented which suggests how A463T and T541A alter the functional structure of WT Towne IEP86. A hydrophobic core containing A463 is predicted to be responsible for the functional integrity of the carboxy-terminal region of IEP86 between amino acids 344 and 579.
Figures


References
-
- Baracchini, E., E. Glezer, K. Fish, R. M. Stenberg, J. A. Nelson, and P. Ghazal. 1992. An isoform variant of the cytomegalovirus immediate-early auto repressor functions as a transcriptional activator. Virology 188:518-529. - PubMed
-
- Boyle, W. J., P. van der Geer, and T. Hunter. 1991. Phosphopeptide mapping and phosphoamino acid analysis by two-dimensional separation on thin-layer cellulose plates. Methods Enzymol. 201:110-149. - PubMed
-
- Brooks, B. R., R. E. Bruccoleri, B. D. Olafson, D. J. States, S. Swaminathan, and M. Karplus. 1983. CHARMM: a program for macromolecular energy, minimization and dynamics calculations. J. Comput. Chem. 4:187-217.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
