

Membrane association of VP22, a herpes simplex virus type 1 tegument protein
- PMID: 12663795
- PMCID: PMC152156
- DOI: 10.1128/jvi.77.8.4888-4898.2003
Membrane association of VP22, a herpes simplex virus type 1 tegument protein
Abstract
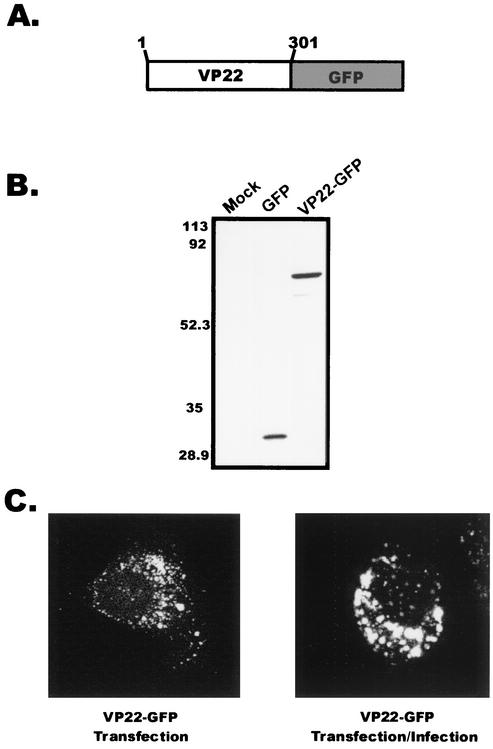
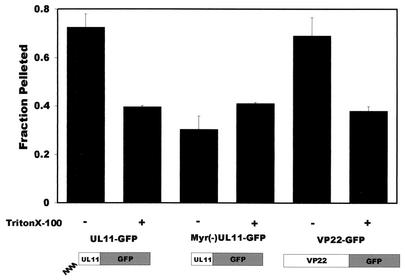
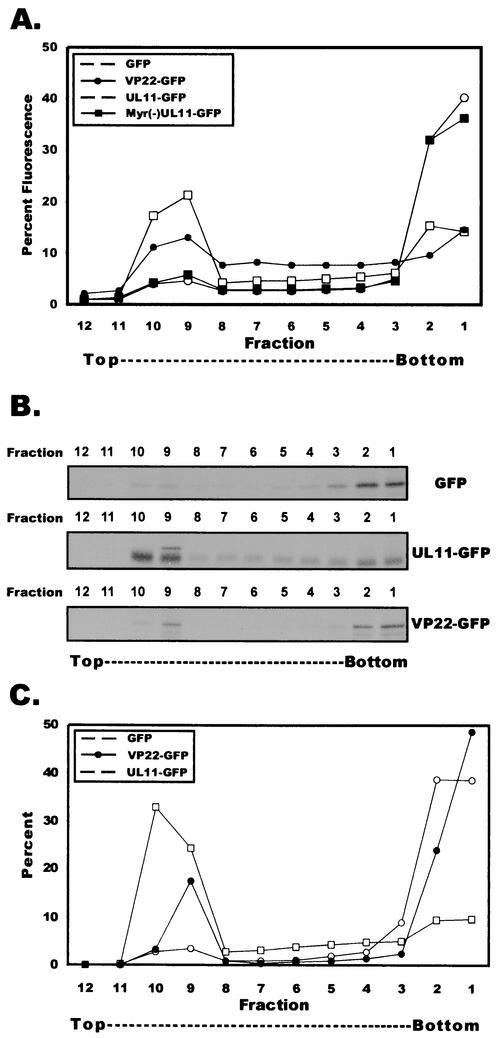
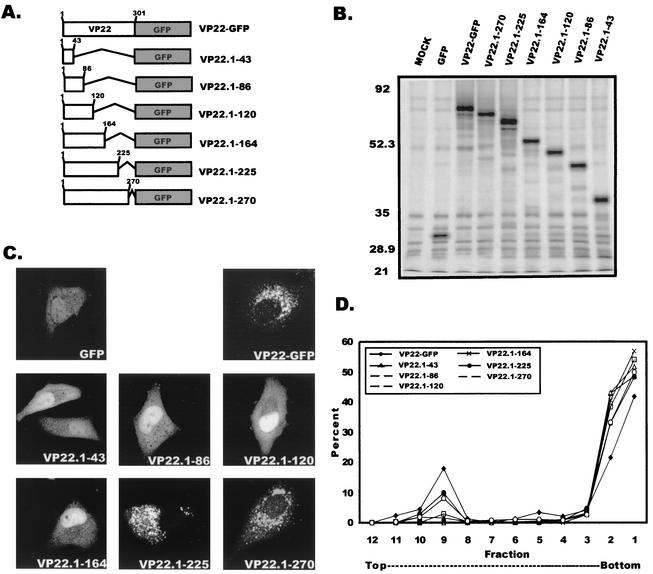
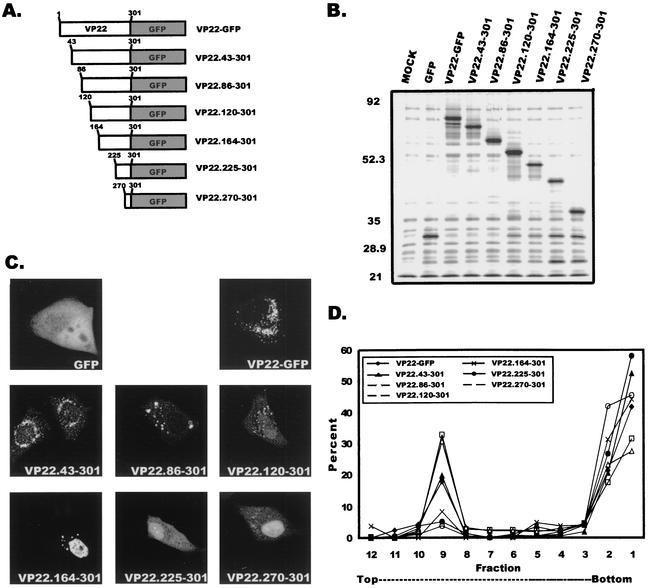
Tegument proteins of herpes simplex virus type 1 (HSV-1) are hypothesized to contain the functional information required for the budding or envelopment process proposed to occur at cytoplasmic compartments of the host cell. One of the most abundant tegument proteins of HSV-1 is the U(L)49 gene product, VP22, a 38-kDa protein of unknown function. To study its subcellular localization, a VP22-green fluorescent protein chimera was expressed in transfected human melanoma (A7) cells. In the absence of other HSV-1 proteins, VP22 localizes to acidic compartments of the cell that may include the trans-Golgi network (TGN), suggesting that this protein is membrane associated. Membrane pelleting and membrane flotation assays confirmed that VP22 partitions with the cellular membrane fraction. Through truncation mutagenesis, we determined that the membrane association of VP22 is a property attributed to amino acids 120 to 225 of this 301-amino-acid protein. The above results demonstrate that VP22 contains specific information required for targeting to membranes of acidic compartments of the cell which may be derived from the TGN, suggesting a potential role for VP22 during tegumentation and/or final envelopment.
Figures


References
-
- Aints, A., H. Guven, G. Gahrton, C. I. Smith, and M. S. Dilber. 2001. Mapping of herpes simplex virus-1 VP22 functional domains for inter- and subcellular protein targeting. Gene Ther. 8:1051-1056. - PubMed
-
- Alconada, A., U. Bauer, L. Baudoux, J. Piette, and B. Hoflack. 1998. Intracellular transport of the glycoproteins gE and gI of the varicella-zoster virus. gE accelerates the maturation of gI and determines its accumulation in the trans-Golgi network. J. Biol. Chem. 273:13430-13436. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
