

A yeast homologue of Hsp70, Ssa1p, regulates turnover of the MFA2 transcript through its AU-rich 3' untranslated region
- PMID: 12665566
- PMCID: PMC152564
- DOI: 10.1128/MCB.23.8.2623-2632.2003
A yeast homologue of Hsp70, Ssa1p, regulates turnover of the MFA2 transcript through its AU-rich 3' untranslated region
Abstract
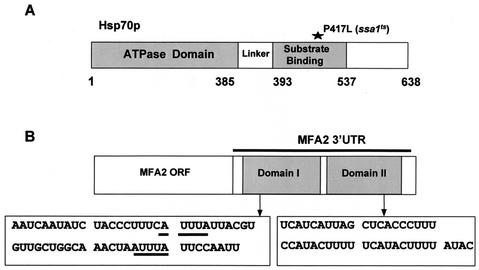
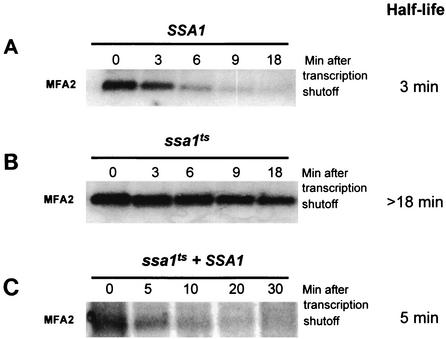
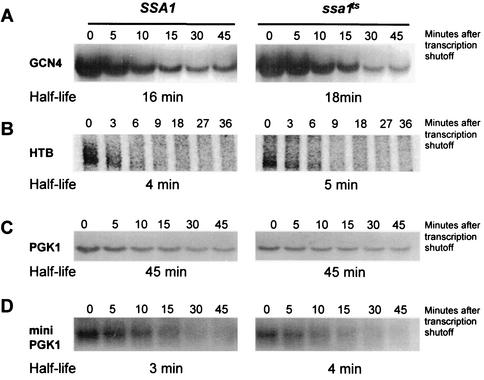
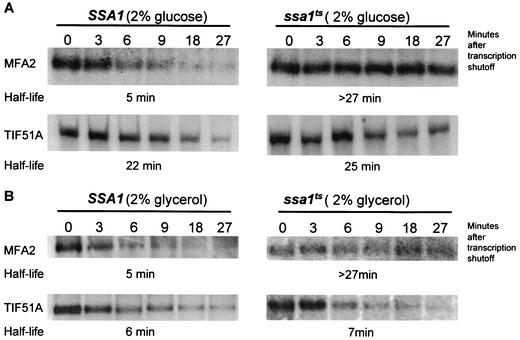
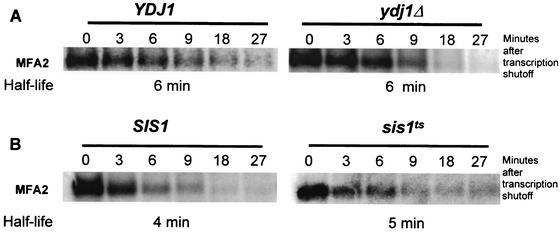
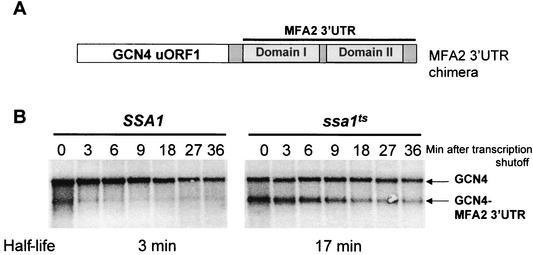
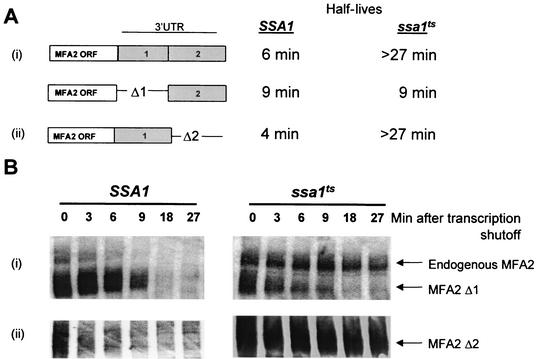
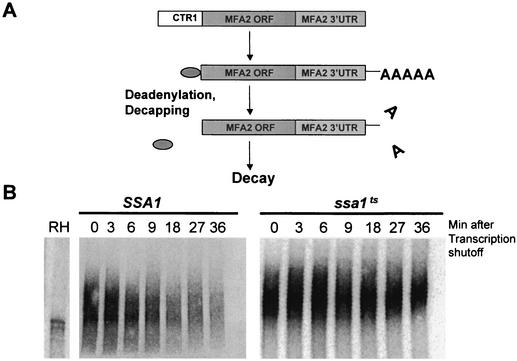
Many eukaryotic mRNAs exhibit regulated decay in response to cellular signals. AU-rich elements (AREs) identified in the 3' untranslated region (3'-UTR) of several such mRNAs play a critical role in controlling the half-lives of these transcripts. The yeast ARE-containing mRNA, MFA2, has been studied extensively and is degraded by a deadenylation-dependent mechanism. However, the trans-acting factors that promote the rapid decay of MFA2 have not been identified. Our results suggest that the chaperone protein Hsp70, encoded by the SSA family of genes, is involved in modulating MFA2 mRNA decay. MFA2 is specifically stabilized in a strain bearing a temperature-sensitive mutation in the SSA1 gene. Furthermore, an AU-rich region within the 3'-UTR of the message is both necessary and sufficient to confer this regulation. Stabilization occurs as a result of slower deadenylation in the ssa1(ts) strain, suggesting that Hsp70 is required for activation of the turnover pathway.
Figures
References
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1992. Current protocols in molecular biology, vol. 2, p. 1-13. John Wiley and Sons, New York, N.Y.
-
- Beere, H. M., and D. R. Green. 2001. Stress management—heat shock protein-70 and the regulation of apoptosis. Trends Cell Biol. 11:6-10. - PubMed
-
- Bernstein, P., and J. Ross. 1989. Poly(A), poly(A) binding protein and the regulation of mRNA stability. Trends Biochem. Sci. 14:373-377. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials