

Upregulation of HMG1 leads to melanoma inhibitory activity expression in malignant melanoma cells and contributes to their malignancy phenotype
- PMID: 12665595
- PMCID: PMC152547
- DOI: 10.1128/MCB.23.8.2991-2998.2003
Upregulation of HMG1 leads to melanoma inhibitory activity expression in malignant melanoma cells and contributes to their malignancy phenotype
Abstract
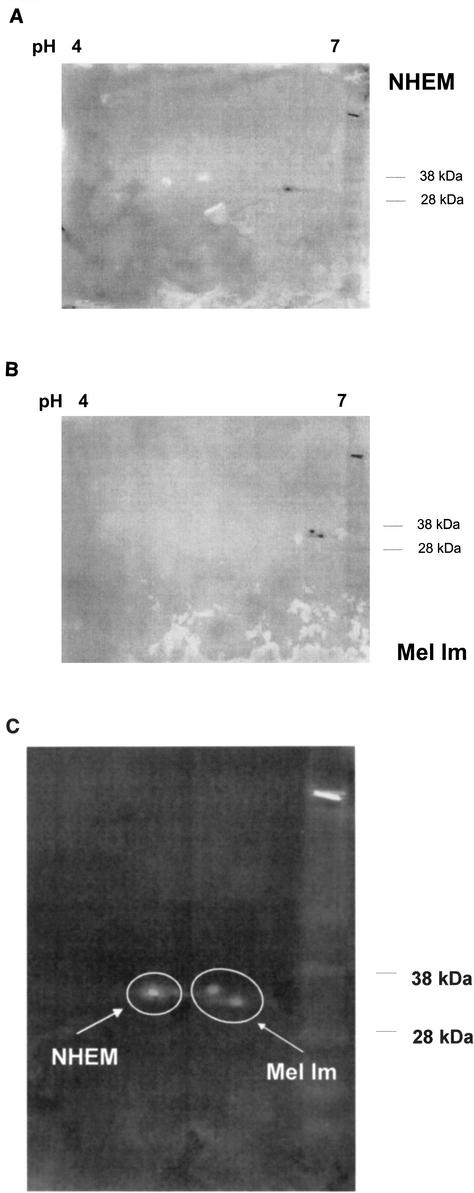
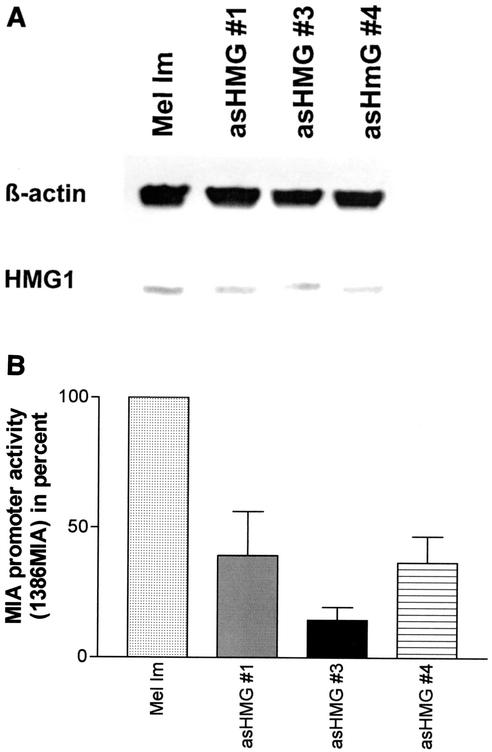
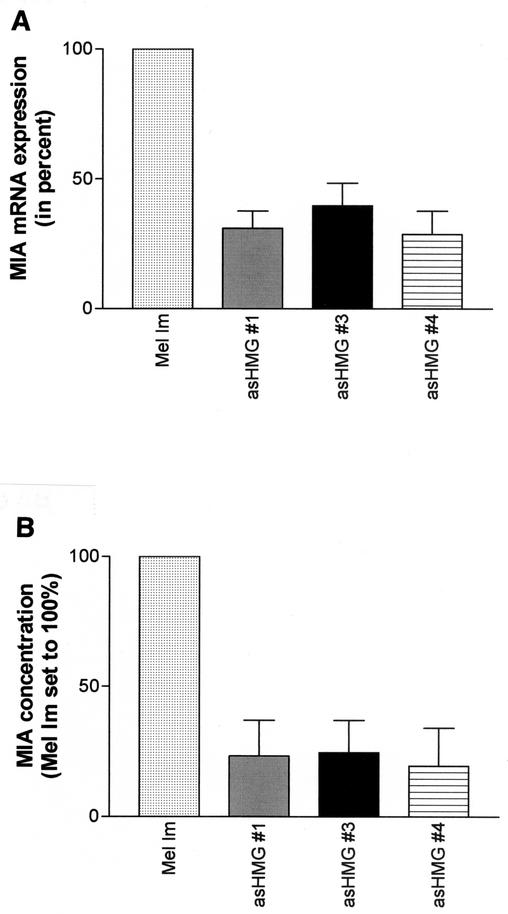
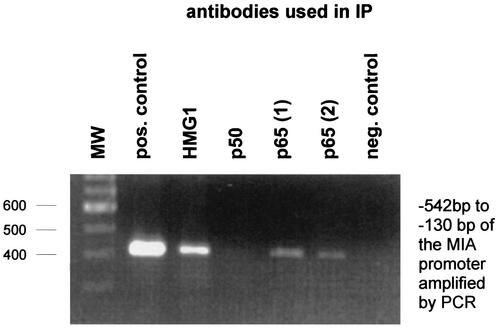
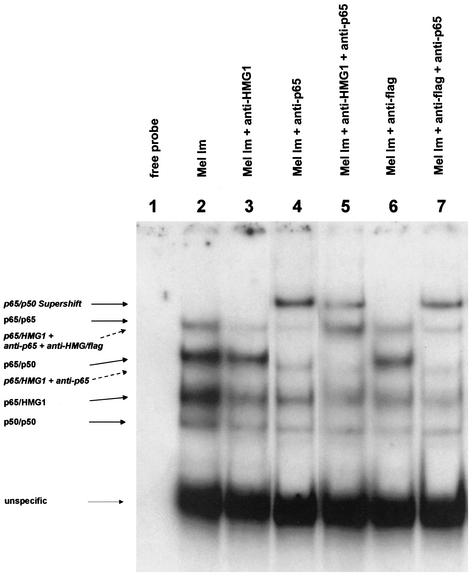
Malignant transformation of melanocytes to melanoma cells closely parallels activation of melanoma inhibitory activity (MIA) expression. We have previously shown that upregulation of MIA occurs on a transcriptional level and involves the highly conserved region (HCR) promoter element. We further observed that the HCR element interacts with the melanoma-associated transcription factor (MATF) and thereby confers strong promoter activation. In this study we identify the peptide sequence of MATF and show that it is identical with the transcription factor HMG1. HMG1 was upregulated in malignant melanoma cells and further activated by hypophosphorylation. Stable antisense-HMG1 expression in melanoma cells led to the reduction of MIA promoter activity and protein expression, indicating that HMG1 is a potent regulator of MIA expression. Interestingly, chromatin immunoprecipitation and electrophoretic mobility shift experiments indicated that HMG1 and the NF-kappa B family member p65 both interact and bind to the HCR promoter element. In summary, our study proves HMG1 and p65 to be important factors in MIA regulation and melanoma progression.
Figures




References
-
- Blesch, A., A. K. Bosserhoff, R. Apfel, C. Behl, B. Hessdoerfer, A. Schmitt, P. Jachimczak, F. Lottspeich, R. Buettner, and U. Bogdahn. 1994. Cloning of a novel malignant melanoma-derived growth-regulatory protein, MIA. Cancer Res. 54:5695-5701. - PubMed
-
- Boonyaratanakornkit, V., V. Melvin, P. Prendergast, M. Altmann, L. Ronfani, M. E. Bianchi, L. Taraseviciene, S. K. Nordeen, E. A. Allegretto, and D. P. Edwards. 1998. High-mobility group chromatin proteins 1 and 2 functionally interact with steroid hormone receptors to enhance their DNA binding in vitro and transcriptional activity in mammalian cells. Mol. Cell. Biol. 18:4471-4487. - PMC - PubMed
-
- Bosserhoff, A. K., A. Hauschild, R. Hein, D. Schadendorf, E. Stockflesh, T. Bogenrieder, M. Landthaler, R. Buettner, and W. Stolz. 2000. Elevated MIA serum levels are of relevance for management of metastasized malignant melanomas: results of a German multicenter study. J. Investig. Dermatol. 114:395-396. - PubMed
-
- Bosserhoff, A. K., R. Hein, U. Bogdahn, and R. Buettner. 1996. Structure and promoter analysis of the gene encoding the human melanoma-inhibiting protein MIA. J. Biol. Chem. 271:490-495. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical