

Chloride currents in acutely isolated Xenopus retinal pigment epithelial cells
- PMID: 12665603
- PMCID: PMC2342951
- DOI: 10.1113/jphysiol.2003.040428
Chloride currents in acutely isolated Xenopus retinal pigment epithelial cells
Abstract
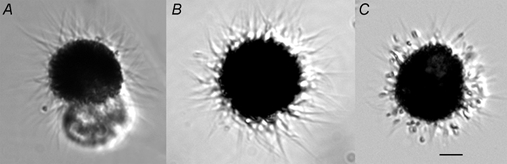
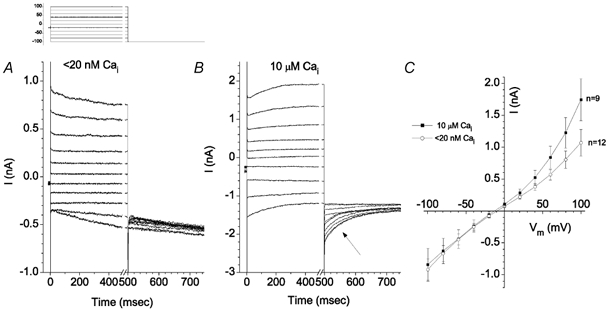
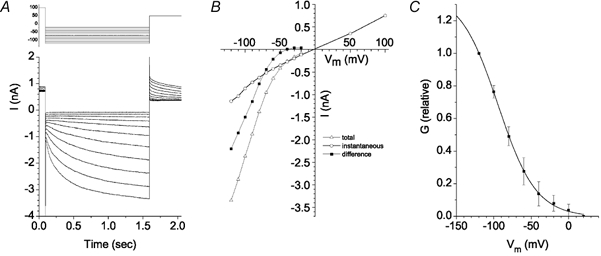
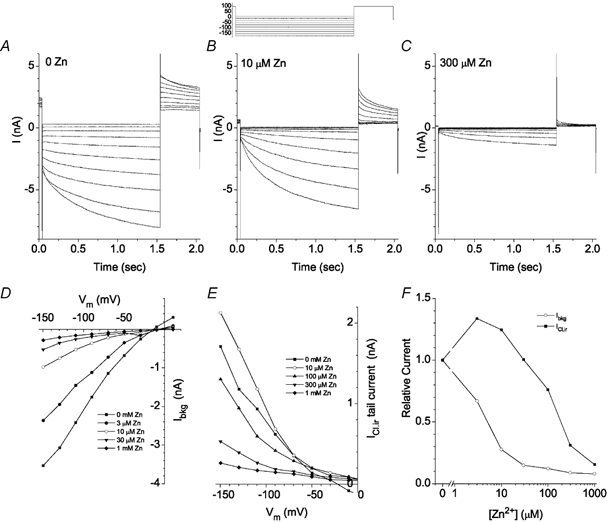
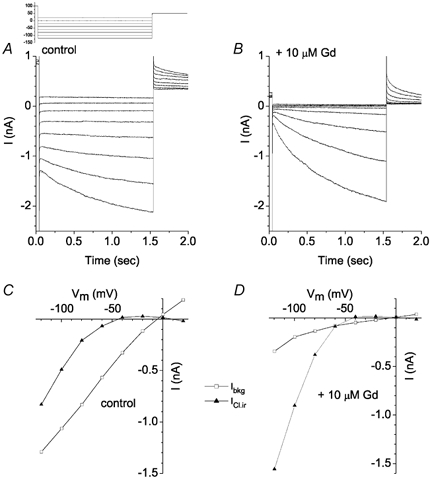
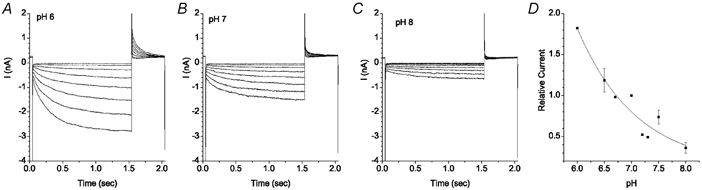
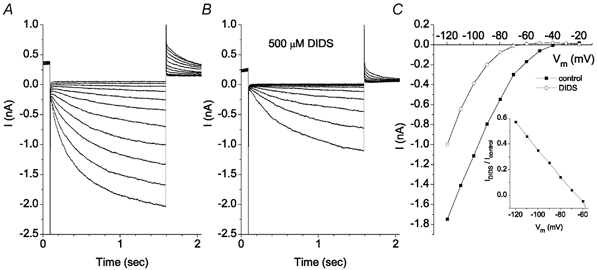
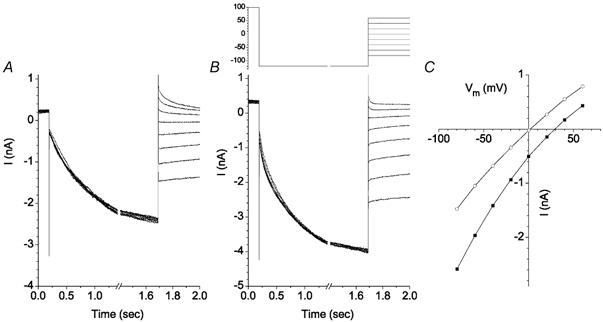
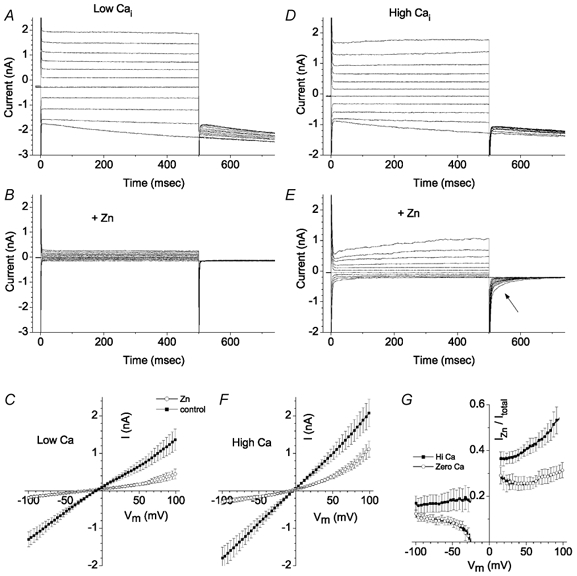
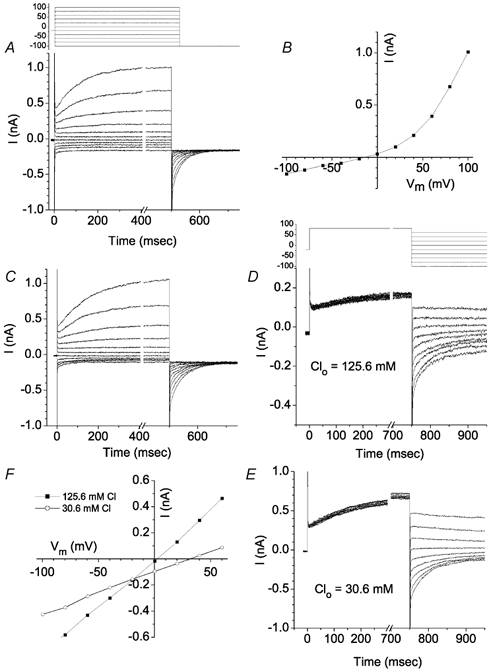
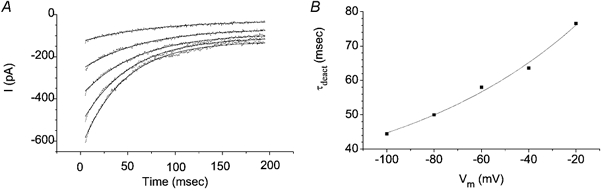
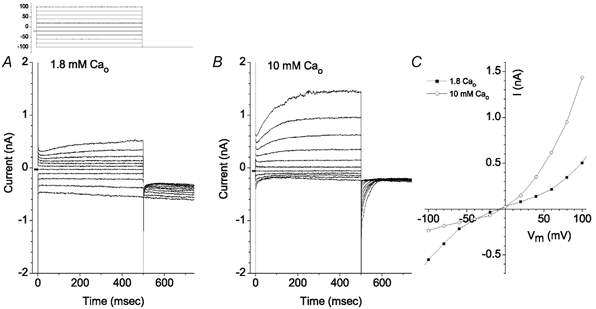
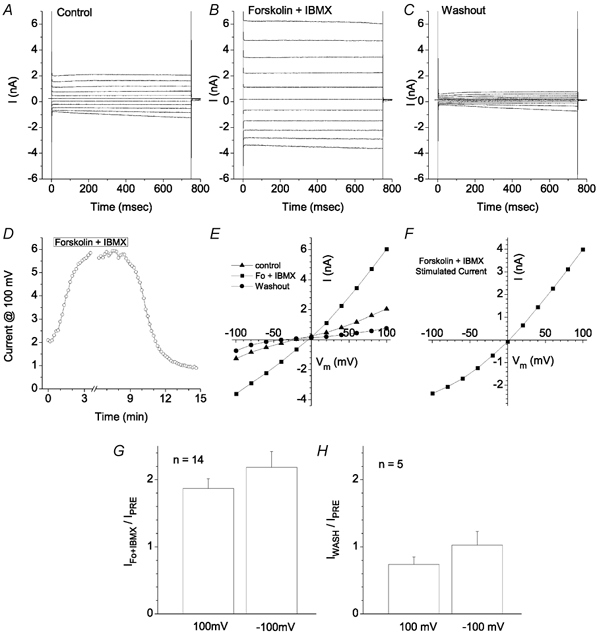
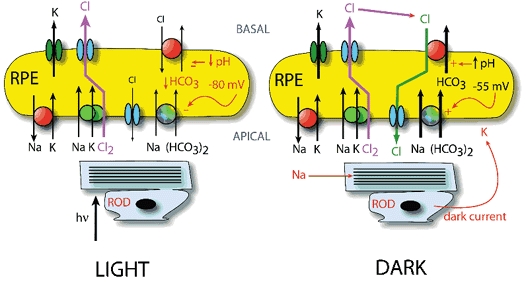
The retinal pigment epithelium (RPE) regulates the ionic composition of the fluid surrounding the photoreceptors by transport mechanisms that utilize Cl- channels. Cl- currents in RPE cells, however, remain incompletely characterized. The purpose of this study was to identify the Cl- currents in acutely isolated Xenopus RPE cells using whole-cell patch clamp. We describe three different Cl- currents. (1) An inwardly rectifying Cl- current, ICl,ir, activates slowly with hyperpolarization (tauact = ~1 s at -80 mV, V1/2= -94 +/- 3 mV), is blocked by Zn2+ (IC50 =185 microM), is stimulated by acid (ICl,ir is 5 times larger at pH 6 than pH 8), and is blocked by DIDS in a voltage-dependent manner. ICl,ir closely resembles cloned ClC-2currents. (2) An outwardly rectifying Cl- current, ICl,Ca, is stimulated by elevated cytosolic free [Ca2+]. With 1 microM free [Ca2+]i in the patch pipette, ICl,Ca activates slowly with depolarization (tauact =325 ms at 100 mV) and deactivates upon hyperpolarization. ICl,Ca is not blocked by 1 mM Zn2+ or 10 microM Gd3+ but is blocked by DIDS. High extracellular [Ca2+] (10 mM) also activates ICl,Ca. (3) A non-rectifying current is activated by elevation of cytoplasmic cAMP with forskolin and IBMX. In addition to these three Cl- currents, Xenopus RPE cells exhibit a non-selective background current (Ibkg) which has a linear I-V relationship and is voltage insensitive. This current is blocked by Zn2+ (IC50 of 5.3 microM) or 10 microM Gd3+. This description provides new insights into the physiology of Cl- channels involved in salt and fluid transport by the retinal pigment epithelium.
Figures
References
-
- Besharse JC, Defoe D. The role of retinal pigment epithelium in photoreceptor membrane turnover. In: Marmor MF, Wolfensberger TJ, editors. The Retinal Pigment Epithelium: Function and Disease. New York: Oxford University Press; 1998. pp. 152–172.
-
- Botchkin LM, Matthews G. Chloride current activated by swelling in retinal pigment epithelium cells. Am J Physiol. 1993;265:C1037–1045. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous