

Herpes simplex virus 1 activates cdc2 to recruit topoisomerase II alpha for post-DNA synthesis expression of late genes
- PMID: 12665617
- PMCID: PMC153640
- DOI: 10.1073/pnas.0730735100
Herpes simplex virus 1 activates cdc2 to recruit topoisomerase II alpha for post-DNA synthesis expression of late genes
Abstract
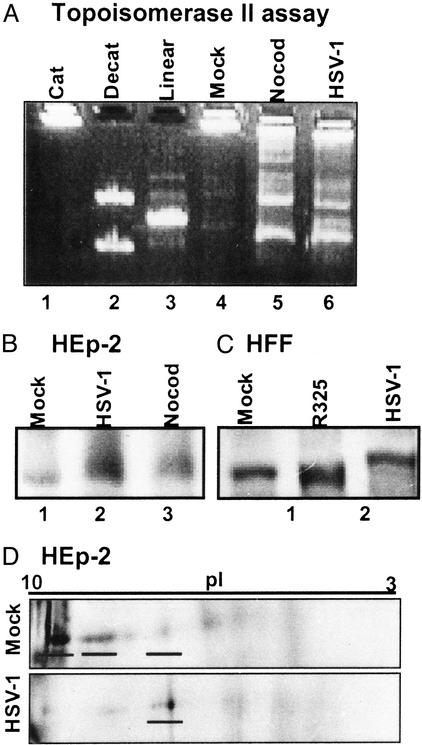
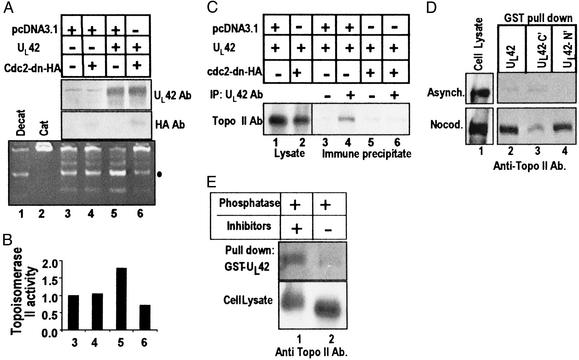
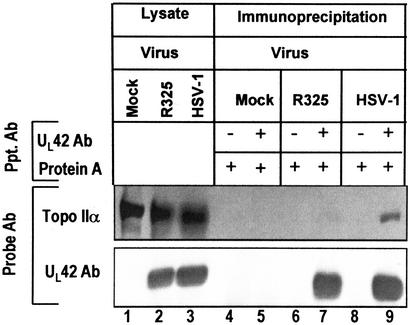
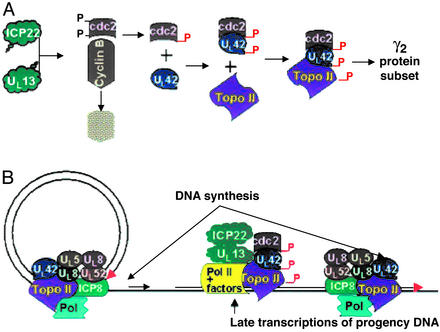
A subset (gamma(2)) of late herpes simplex virus 1 genes depends on viral DNA synthesis for its expression. For optimal expression, a small number of these genes, exemplified by U(S)11, also requires two viral proteins, the alpha protein infected cell protein (ICP) 22 and the protein kinase U(L)13. Earlier we showed that U(L)13 and ICP22 mediate the stabilization of cdc2 and the replacement of its cellular partner, cyclin B, with the viral DNA polymerase processivity factor U(L)42. Here we report that cdc2 and its new partner, U(L)42, bind a phosphorylated form of topoisomerase II alpha. The posttranslational modification of topoisomerase II alpha and its interaction with cdc2-U(L)42 proteins depend on ICP22 in infected cells. Although topoisomerase II is required for viral DNA synthesis, ICP22 is not, indicating a second function for topoisomerase II alpha. The intricate manner in which the virus recruits topoisomerase II alpha for post-DNA synthesis expression of viral genes suggests that topoisomerase II alpha also is required for untangling concatemeric DNA progeny for optimal transcription of late genes.
Figures
References
-
- Roizman B, Sears A E. In: Fields Virology. Fields B N, Knipe D M, Howley P, Chanock R M, Hirsch M S, Melnick J L, Monath T P, Roizman B, editors. New York: Lippincott-Raven; 1996. pp. 2231–2296.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
