

Combinatorial modification of multiple lignin traits in trees through multigene cotransformation
- PMID: 12668766
- PMCID: PMC153659
- DOI: 10.1073/pnas.0831166100
Combinatorial modification of multiple lignin traits in trees through multigene cotransformation
Abstract
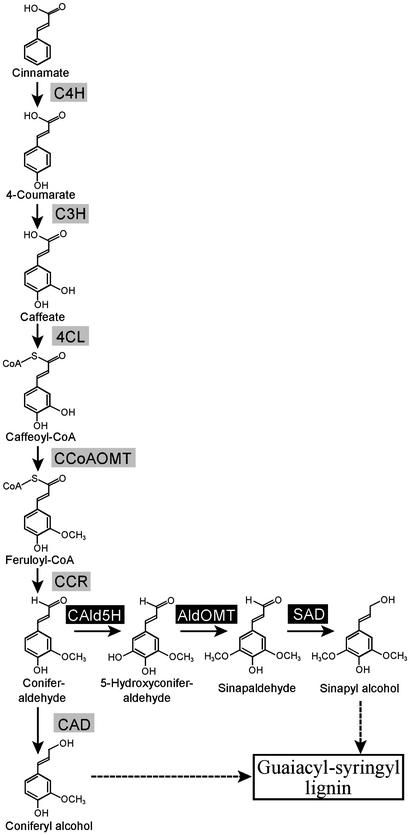
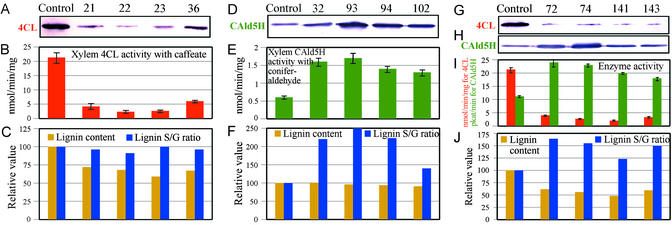
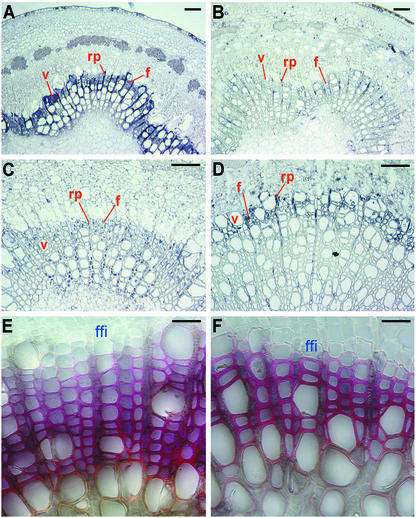
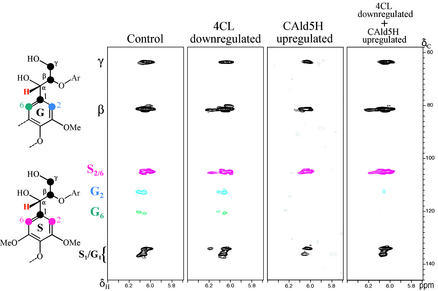
Lignin quantity and reactivity [which is associated with its syringyl/guaiacyl (S/G) constituent ratio] are two major barriers to wood-pulp production. To verify our contention that these traits are regulated by distinct monolignol biosynthesis genes, encoding 4-coumarate-CoA ligase (4CL) and coniferaldehyde 5-hydroxylase (CAld5H), we used Agrobacterium to cotransfer antisense 4CL and sense CAld5H genes into aspen (Populus tremuloides). Trees expressing each one and both of the transgenes were produced with high efficiency. Lignin reduction by as much as 40% with 14% cellulose augmentation was achieved in antisense 4CL plants; S/G-ratio increases as much as 3-fold were observed without lignin quantity change in sense CAld5H plants. Consistent with our contention, these effects were independent but additive, with plants expressing both transgenes having up to 52% less lignin, a 64% higher S/G ratio, and 30% more cellulose. An S/G-ratio increase also accelerated cell maturation in stem secondary xylem, pointing to a role for syringyl lignin moieties in coordinating xylem secondary wall biosynthesis. The results suggest that this multigene cotransfer system should be broadly useful for plant genetic engineering and functional genomics.
Figures
References
-
- Whetten R W, MacKay J J, Sederoff R R. Plant Mol Biol. 1998;49:585–609. - PubMed
-
- Hu W-J, Lung J, Harding S A, Popko J L, Ralph J, Stokke D D, Tsai C-J, Chiang V L. Nat Biotechnol. 1999;17:808–812. - PubMed
-
- Pilate G, Guieny E, Holt K, Petit-Conil M, Lapierre C, Leple J-C, Pollet B, Mila I, Webster E A, Marstorp G G, et al. Nat Biotechnol. 2002;20:607–612. - PubMed
-
- Chiang V L. Nat Biotechnol. 2002;20:557–558. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
