

In vivo analysis of a developmental circuit for direct transcriptional activation and repression in the same cell by a Runx protein
- PMID: 12670867
- PMCID: PMC196027
- DOI: 10.1101/gad.1064803
In vivo analysis of a developmental circuit for direct transcriptional activation and repression in the same cell by a Runx protein
Abstract
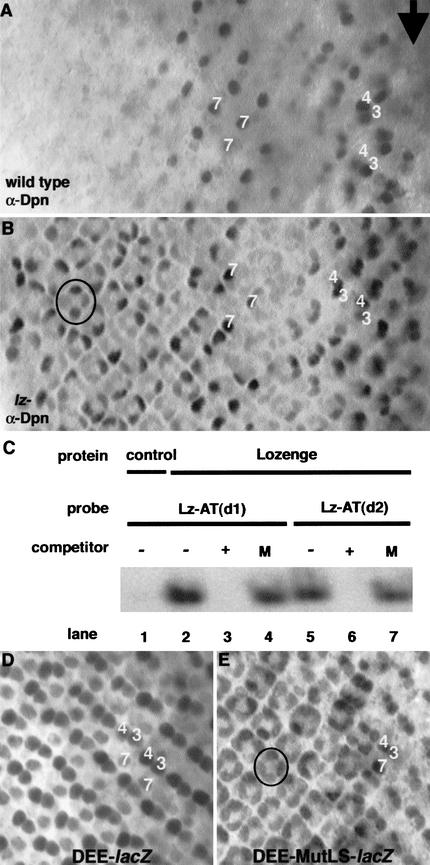
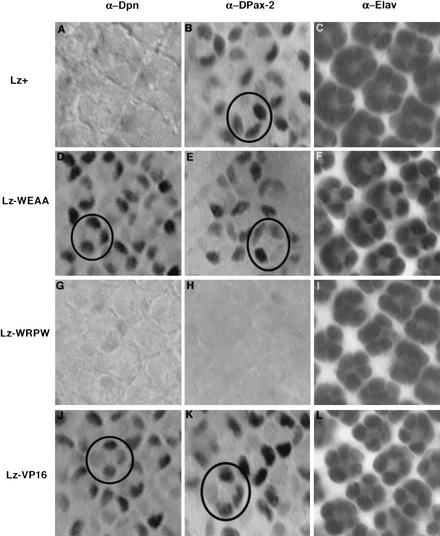
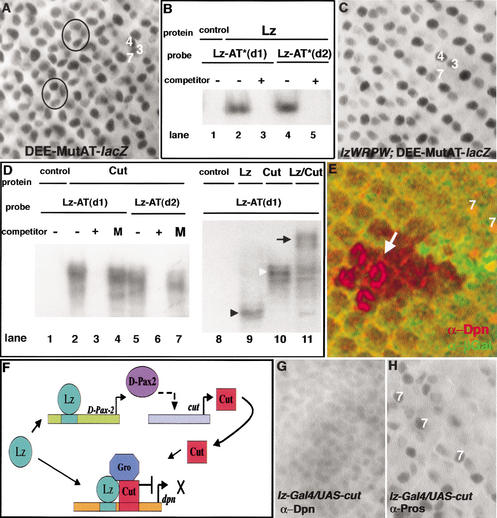
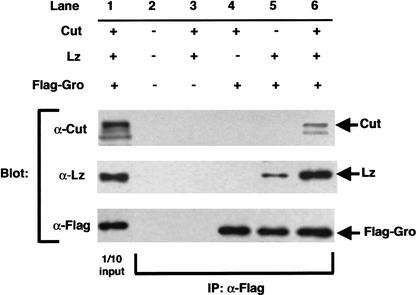
Runx proteins have been implicated in acute myeloid leukemia, cleidocranial dysplasia, and stomach cancer. These proteins control key developmental processes in which they function as both transcriptional activators and repressors. How these opposing regulatory modes can be accomplished in the in vivo context of a cell has not been clear. In this study we use the developing cone cell in the Drosophila visual system to elucidate the mechanism of positive and negative regulation by the Runx protein Lozenge (Lz). We describe a regulatory circuit in which Lz causes transcriptional activation of the homeodomain protein Cut, which can then stabilize a Lz repressor complex in the same cell. Whether a gene is activated or repressed is determined by whether the Lz activator or the repressor complex binds to its upstream sequence. This study provides a mechanistic basis for the dual function of Runx proteins that is likely to be conserved in mammalian systems.
Figures
References
-
- Bier E, Vaessin H, Younger-Shepherd S, Jan LY, Jan YN. deadpan, an essential pan-neural gene in Drosophila, encodes a helix–loop–helix protein similar to the hairy gene product. Genes & Dev. 1992;6:2137–2151. - PubMed
-
- Blochlinger K, Bodmer R, Jack J, Jan LY, Jan YN. Primary structure and expression of a product from cut, a locus involved in specifying sensory organ identity in Drosophila. Nature. 1988;333:629–635. - PubMed
-
- Canon J, Banerjee U. Runt and Lozenge function in Drosophila development. Semin Cell Dev Biol. 2000;11:327–336. - PubMed
-
- Castilla LH, Wijmenga C, Wang Q, Stacy T, Speck NA, Eckhaus M, Marin-Padilla M, Collins FS, Wynshaw-Boris A, Liu PP. Failure of embryonic hematopoiesis and lethal hemorrhages in mouse embryos heterozygous for a knocked-in leukemia gene CBFB-MYH11. Cell. 1996;87:687–696. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases