

Treatment with isotretinoin inhibits lipofuscin accumulation in a mouse model of recessive Stargardt's macular degeneration
- PMID: 12671074
- PMCID: PMC153626
- DOI: 10.1073/pnas.0737855100
Treatment with isotretinoin inhibits lipofuscin accumulation in a mouse model of recessive Stargardt's macular degeneration
Abstract
Recessive Stargardt's macular degeneration is an inherited blinding disease of children caused by mutations in the ABCR gene. The primary pathologic defect in Stargardt's disease is accumulation of toxic lipofuscin pigments such as N-retinylidene-N-retinylethanolamine (A2E) in cells of the retinal pigment epithelium. This accumulation appears to be responsible for the photoreceptor death and severe visual loss in Stargardt's patients. Here, we tested a therapeutic strategy to inhibit lipofuscin accumulation in a mouse model of recessive Stargardt's disease. Isotretinoin (Accutane) has been shown to slow the synthesis of 11-cis-retinaldehyde and regeneration of rhodopsin by inhibiting 11-cis-retinol dehydrogenase in the visual cycle. Light activation of rhodopsin results in its release of all-trans-retinaldehyde, which constitutes the first reactant in A2E biosynthesis. Accordingly, we tested the effects of isotretinoin on lipofuscin accumulation in abcr(-/-) knockout mice. Isotretinoin blocked the formation of A2E biochemically and the accumulation of lipofuscin pigments by electron microscopy. We observed no significant visual loss in treated abcr(-/-) mice by electroretinography. Isotretinoin also blocked the slower, age-dependent accumulation of lipofuscin in wild-type mice. These results corroborate the proposed mechanism of A2E biogenesis. Further, they suggest that treatment with isotretinoin may inhibit lipofuscin accumulation and thus delay the onset of visual loss in Stargardt's patients. Finally, the results suggest that isotretinoin may be an effective treatment for other forms of retinal or macular degeneration associated with lipofuscin accumulation.
Figures




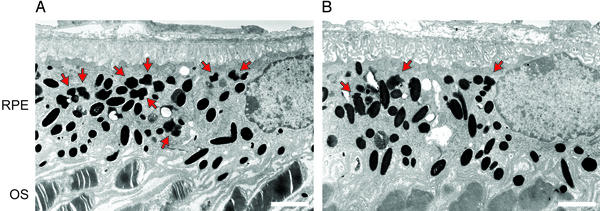
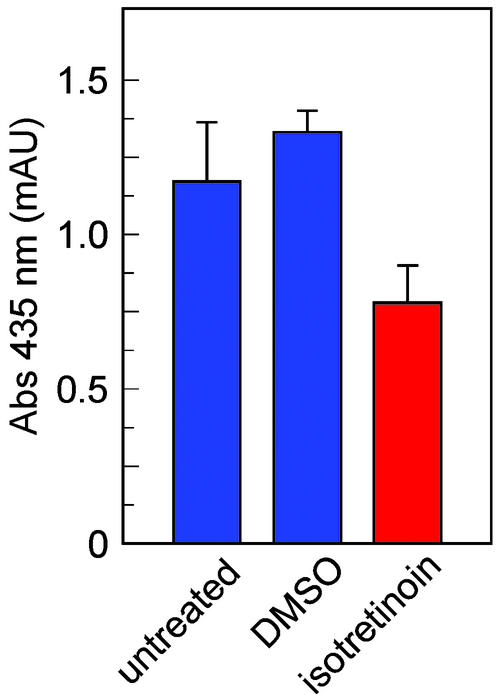
Comment in
-
Therapy for macular degeneration: insights from acne.Proc Natl Acad Sci U S A. 2003 Apr 15;100(8):4353-4. doi: 10.1073/pnas.1031478100. Epub 2003 Apr 7. Proc Natl Acad Sci U S A. 2003. PMID: 12682280 Free PMC article. No abstract available.
References
-
- Leibowitz H M, Krueger D E, Maunder L R, Milton R C, Kini M M, Kahn H A, Nickerson R J, Pool J, Colton T L, Ganley J P, et al. Surv Ophthalmol. 1980;24:335–610. - PubMed
-
- Klein R, Klein B E, Linton K L P. Ophthalmology. 1992;99:933–943. - PubMed
-
- Lee B L, Heckenlively J R. In: Retina-Vitreous-Macula. Guyer D R, Yannuzzi L A, Chang S, Shields J A, Green W R, editors. Philadelphia: Saunders; 1999. pp. 978–988.
-
- Blacharski P A. Fundus flavimaculutus. New York: Raven; 1988.
-
- Eagle R C, Jr, Lucier A C, Bernardino V B, Jr, Yanoff M. Ophthalmology. 1980;87:1189–1200. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
