

Members of the Arabidopsis dynamin-like gene family, ADL1, are essential for plant cytokinesis and polarized cell growth
- PMID: 12671086
- PMCID: PMC524700
- DOI: 10.1105/tpc.009670
Members of the Arabidopsis dynamin-like gene family, ADL1, are essential for plant cytokinesis and polarized cell growth
Abstract
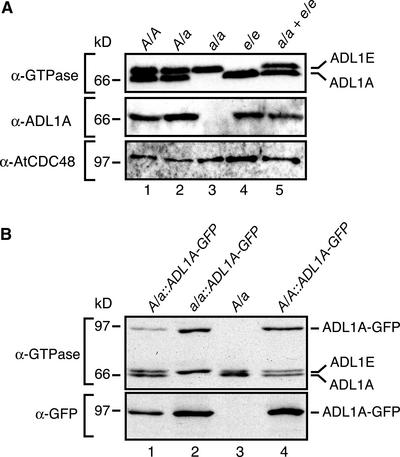
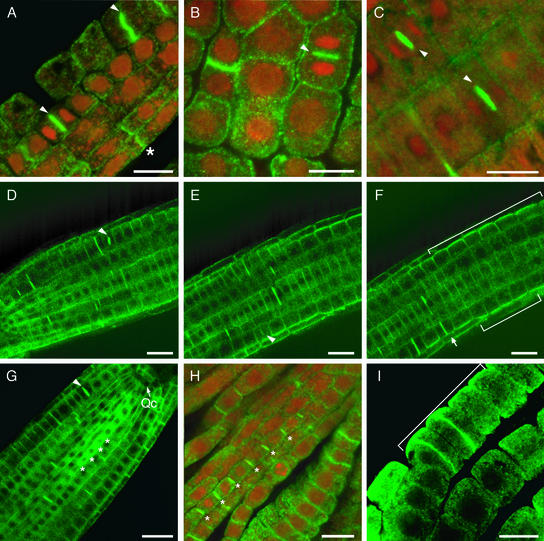
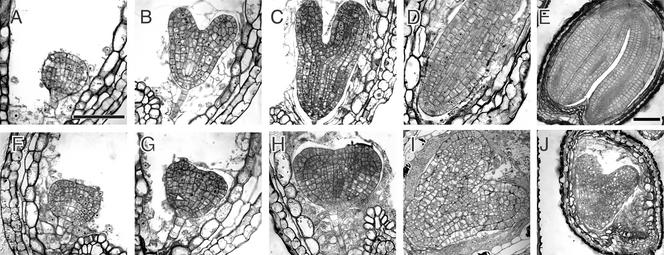
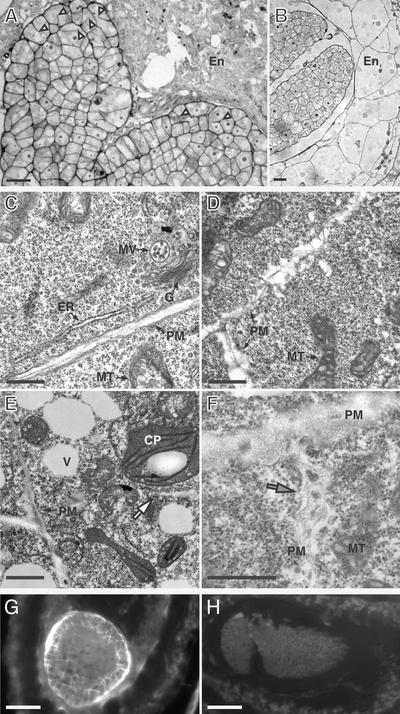
Polarized membrane trafficking during plant cytokinesis and cell expansion are critical for plant morphogenesis, yet very little is known about the molecular mechanisms that guide this process. Dynamin and dynamin-related proteins are large GTP binding proteins that are involved in membrane trafficking. Here, we show that two functionally redundant members of the Arabidopsis dynamin-related protein family, ADL1A and ADL1E, are essential for polar cell expansion and cell plate biogenesis. adl1A-2 adl1E-1 double mutants show defects in cell plate assembly, cell wall formation, and plasma membrane recycling. Using a functional green fluorescent protein fusion protein, we show that the distribution of ADL1A is dynamic and that the protein is localized asymmetrically to the plasma membrane of newly formed and mature root cells. We propose that ADL1-mediated membrane recycling is essential for plasma membrane formation and maintenance in plants.
Figures
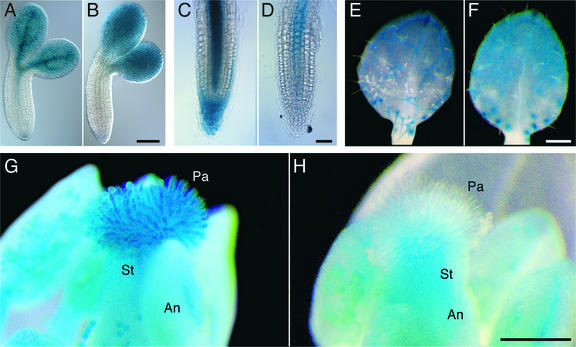
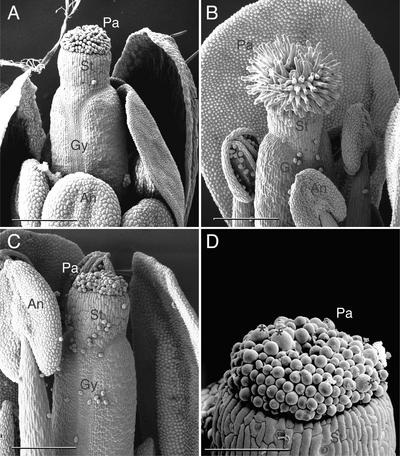
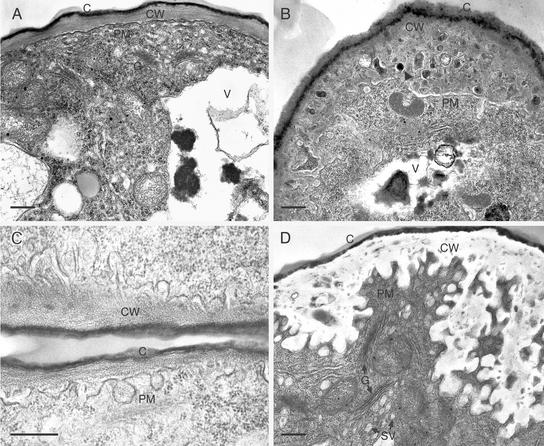

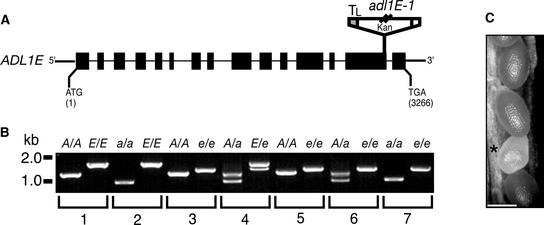
References
-
- Assaad, F.F., Mayer, U., Wanner, G., and Jürgens, G. (1996). The KEULE gene is involved in cytokinesis in Arabidopsis. Mol. Gen. Genet. 253, 267–277. - PubMed
-
- Bednarek, S.Y., and Falbel, T.G. (2002). Membrane trafficking during plant cytokinesis. Traffic 3, 621–629. - PubMed
-
- Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation. Plant J. 16, 735–743. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
