

Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress
- PMID: 12671093
- PMCID: PMC152344
- DOI: 10.1105/tpc.010405
Zeaxanthin accumulation in the absence of a functional xanthophyll cycle protects Chlamydomonas reinhardtii from photooxidative stress
Abstract
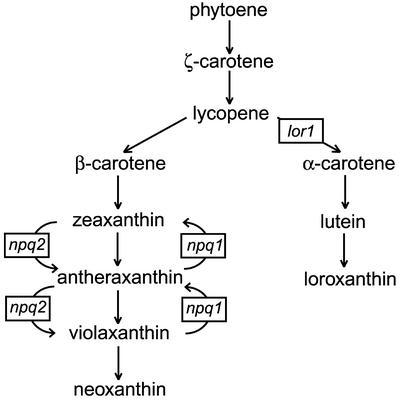
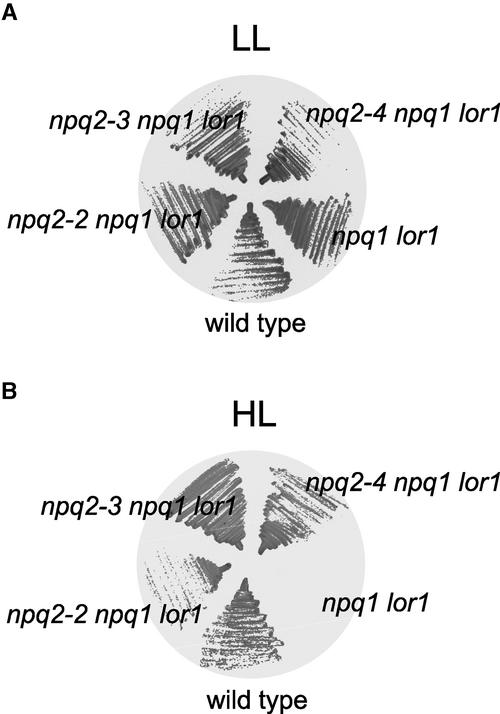
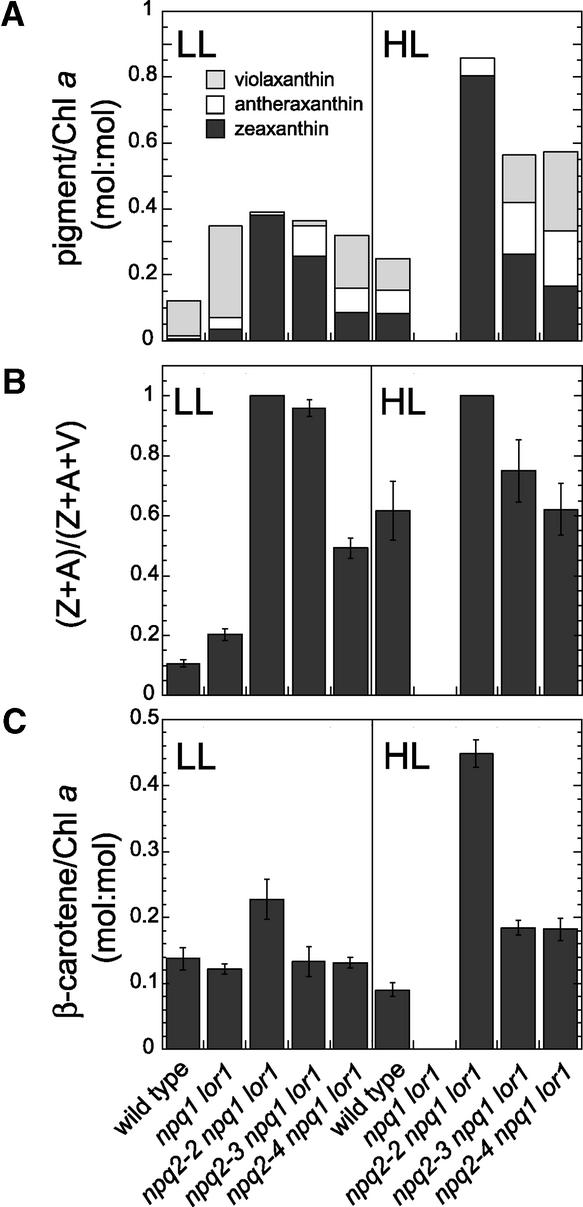
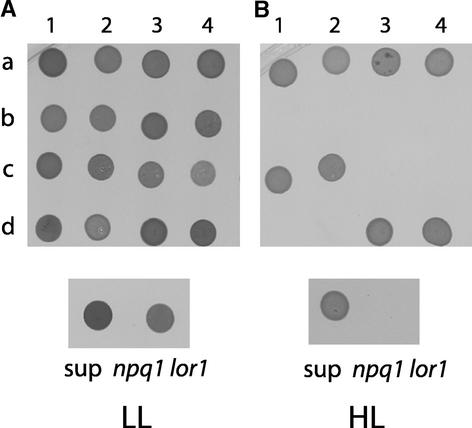
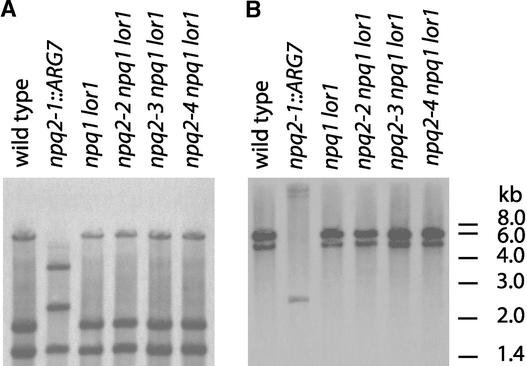
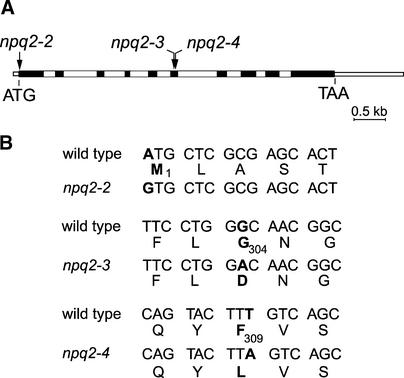
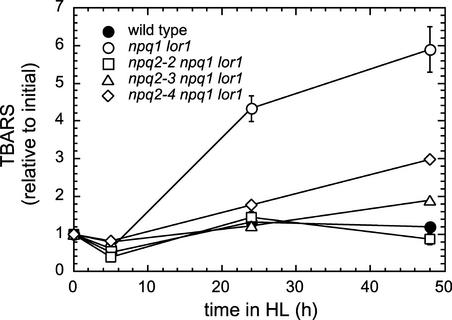
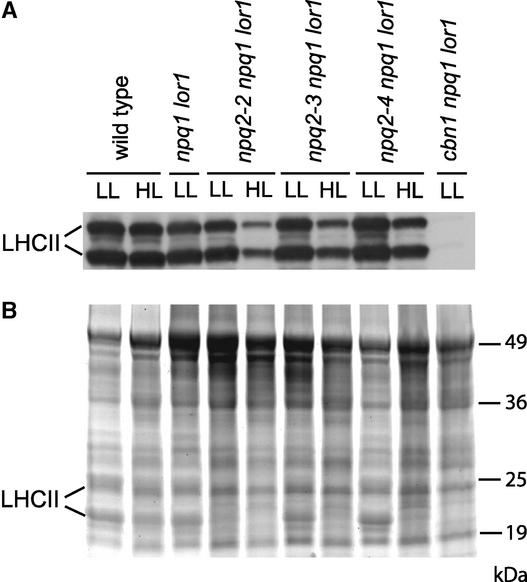
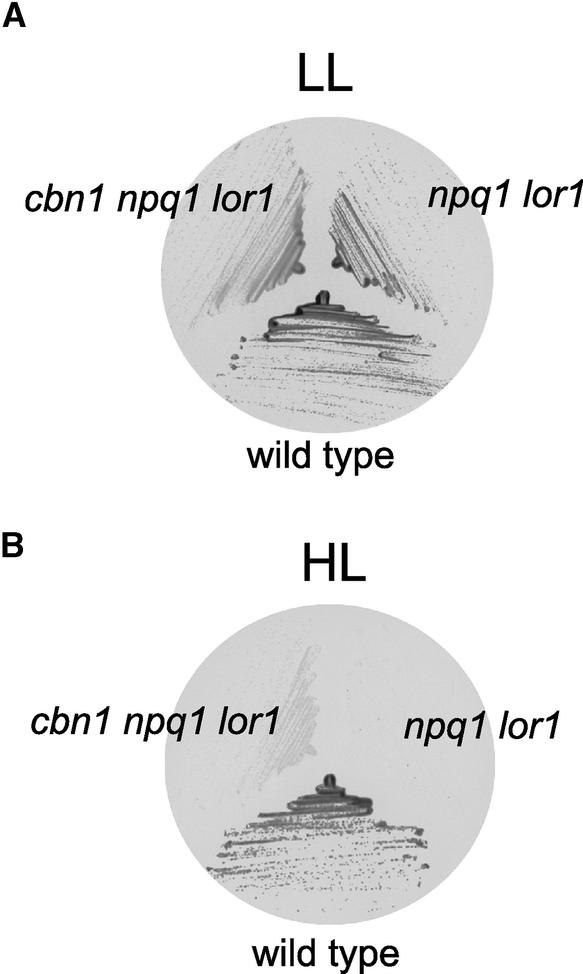
Xanthophylls participate in light harvesting and are essential in protecting the chloroplast from photooxidative damage. To investigate the roles of xanthophylls in photoprotection, we isolated and characterized extragenic suppressors of the npq1 lor1 double mutant of Chlamydomonas reinhardtii, which lacks zeaxanthin and lutein and undergoes irreversible photooxidative bleaching and cell death at moderate to high light intensities. Here, we describe three suppressor strains that carry point mutations in the coding sequence of the zeaxanthin epoxidase gene, resulting in the constitutive accumulation of zeaxanthin in a range of concentrations. The presence of zeaxanthin in these strains was sufficient to prevent photooxidative damage in the npq1 lor1 background. The size of the light-harvesting antenna in the suppressors decreased in high light in a manner that was proportional to the relative content of zeaxanthin, with the strain having the most zeaxanthin showing a severe reduction in levels of the major light-harvesting complex II proteins in high light. We show that the effect of constitutive zeaxanthin on light harvesting is not the main cause of increased photoprotection, because in the absence of zeaxanthin, a strain with a smaller light-harvesting antenna showed only minor protection against photobleaching in high light. Furthermore, the zeaxanthin-accumulating suppressors were able to tolerate higher levels of exogenous reactive oxygen than their parental strain under conditions that did not affect light harvesting. Our results are consistent with an antioxidant role of zeaxanthin in the quenching of singlet oxygen and/or free radicals in the thylakoid membrane in vivo.
Figures


References
-
- Allen, K.D., and Staehelin, L.A. (1994). Polypeptide composition, assembly and phosphorylation patterns of the photosystem II antenna system of Chlamydomonas reinhardtii. Planta 194, 42–54.
-
- Asamizu, E., Miura, K., Kucho, K., Inoue, Y., Fukuzawa, H., Ohyama, K., Nakamura, Y., and Tabata, S. (2000). Generation of expressed sequence tags from low-CO2 and high-CO2 adapted cells of Chlamydomonas reinhardtii. DNA Res. 7, 305–307. - PubMed
-
- Bassi, R., and Caffarri, S. (2000). Lhc proteins and the regulation of photosynthetic light harvesting function by xanthophylls. Photosynth. Res. 64, 243–256. - PubMed
-
- Bassi, R., and Wollman, F.-A. (1991). The chlorophyll-a/b proteins of photosystem II in Chlamydomonas reinhardtii: Isolation, characterization and immunological cross-reactivity to higher-plant polypeptides. Planta 183, 423–433. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
