

Neural, not gonadal, origin of brain sex differences in a gynandromorphic finch
- PMID: 12672961
- PMCID: PMC153648
- DOI: 10.1073/pnas.0636925100
Neural, not gonadal, origin of brain sex differences in a gynandromorphic finch
Abstract
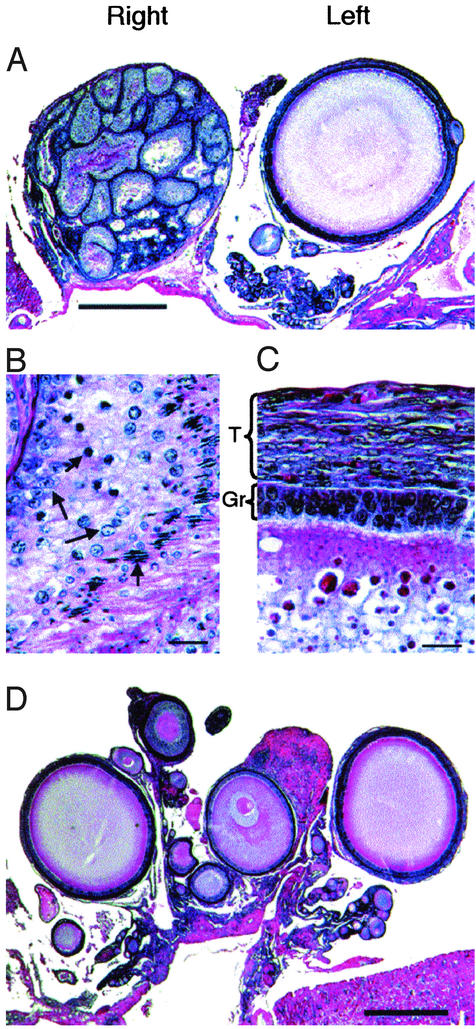
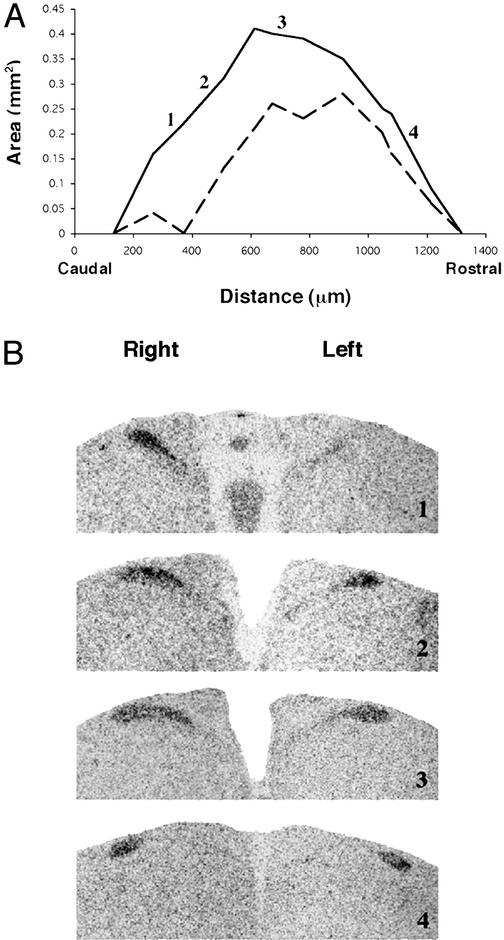
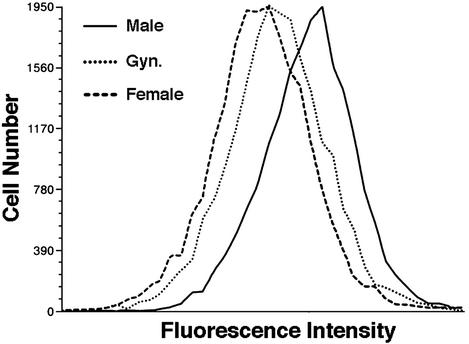
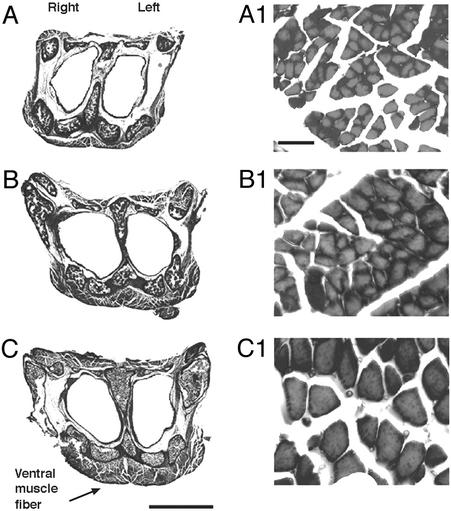
In mammals and birds, sex differences in brain function and disease are thought to derive exclusively from sex differences in gonadal hormone secretions. For example, testosterone in male mammals acts during fetal and neonatal life to cause masculine neural development. However, male and female brain cells also differ in genetic sex; thus, sex chromosome genes acting within cells could contribute to sex differences in cell function. We analyzed the sexual phenotype of the brain of a rare gynandromorphic finch in which the right half of the brain was genetically male and the left half genetically female. The neural song circuit on the right had a more masculine phenotype than that on the left. Because both halves of the brain were exposed to a common gonadal hormone environment, the lateral differences indicate that the genetic sex of brain cells contributes to the process of sexual differentiation. Because both sides of the song circuit were more masculine than that of females, diffusible factors such as hormones of gonadal or neural origin also likely played a role in sexual differentiation.
Figures






References
-
- Arnold A P. In: Hormones, Brain, and Behavior. Pfaff D W, Etgen A, Fahrbach S, Rubin R, editors. Vol. 4. New York: Academic; 2002. pp. 105–135.
-
- Goodfellow P N, Lovell-Badge R. Annu Rev Genet. 1993;27:71–92. - PubMed
-
- Arnold A P, Gorski R A. Annu Rev Neurosci. 1984;7:413–442. - PubMed
-
- Phoenix C, Goy R, Gerall A A, Young W C. Endocrinology. 1959;65:369–382. - PubMed
-
- Nottebohm F, Arnold A P. Science. 1976;194:211–213. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
