

A new rate law describing microbial respiration
- PMID: 12676718
- PMCID: PMC154818
- DOI: 10.1128/AEM.69.4.2340-2348.2003
A new rate law describing microbial respiration
Abstract
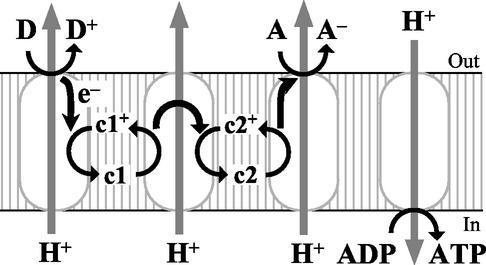
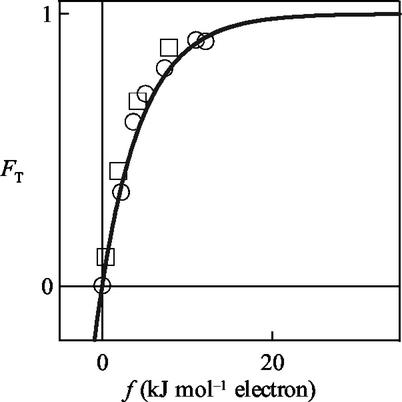
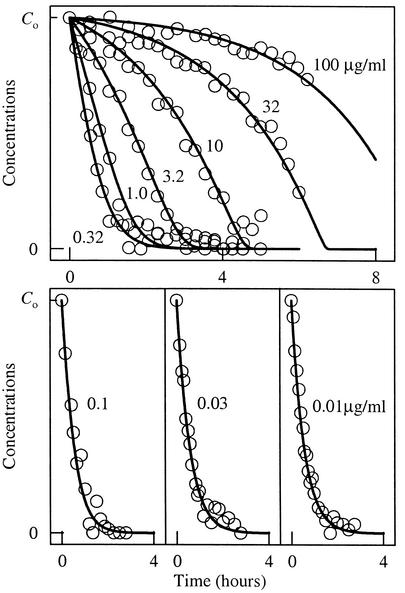
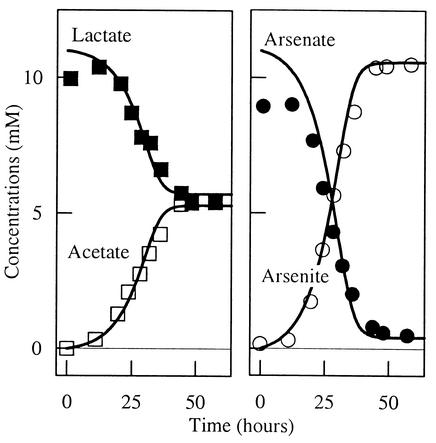
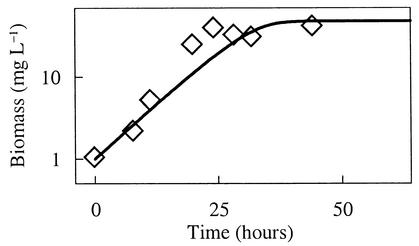
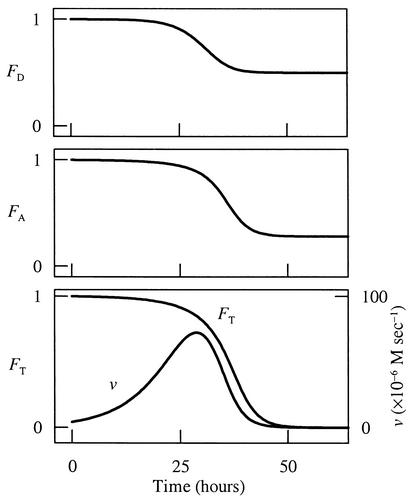
The rate of microbial respiration can be described by a rate law that gives the respiration rate as the product of a rate constant, biomass concentration, and three terms: one describing the kinetics of the electron-donating reaction, one for the kinetics of the electron-accepting reaction, and a thermodynamic term accounting for the energy available in the microbe's environment. The rate law, derived on the basis of chemiosmotic theory and nonlinear thermodynamics, is unique in that it accounts for both forward and reverse fluxes through the electron transport chain. Our analysis demonstrates how a microbe's respiration rate depends on the thermodynamic driving force, i.e., the net difference between the energy available from the environment and energy conserved as ATP. The rate laws commonly applied in microbiology, such as the Monod equation, are specific simplifications of the general law presented. The new rate law is significant because it affords the possibility of extrapolating in a rigorous manner from laboratory experiment to a broad range of natural conditions, including microbial growth where only limited energy is available. The rate law also provides a new explanation of threshold phenomena, which may reflect a thermodynamic equilibrium where the energy released by electron transfer balances that conserved by ADP phosphorylation.
Figures
References
-
- Bae, W., and B. E. Rittmann. 1996. A stuctured model of dual-limitation kinetics. Biotechnol. Bioeng. 49:683-689. - PubMed
-
- Bekins, B. A., E. Warren, and E. M. Godsy. 1998. A comparision of zero-order, first-order, and Monod biotransformation models. Ground Water 36:261-268.
-
- Bethke, C. M. 1996. Geochemical reaction modeling. Oxford University Press, New York, N.Y.
-
- Bethke, C. M. 2002. The geochemist's workbench, release 4.0. University of Illinois, Urbana.
-
- Blum, J. S., A. B. Bindi, J. Buzzelli, J. F. Stolz, and R. S. Oremland. 1998. Bacillus arsenicoselenatis, sp. nov., and Bacillus selenitireducens, sp. nov.: two haloalkaliphiles from Mono Lake, California, that respire oxyanions of selenium and arsenic. Arch. Microbiol. 171:19-30. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
