

Plasma membrane localization and function of the estrogen receptor alpha variant (ER46) in human endothelial cells
- PMID: 12682286
- PMCID: PMC153637
- DOI: 10.1073/pnas.0831079100
Plasma membrane localization and function of the estrogen receptor alpha variant (ER46) in human endothelial cells
Abstract
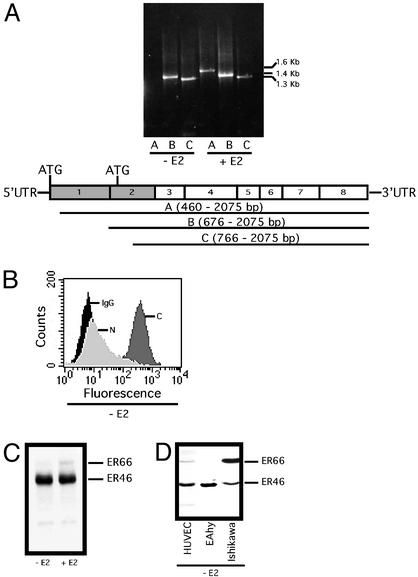
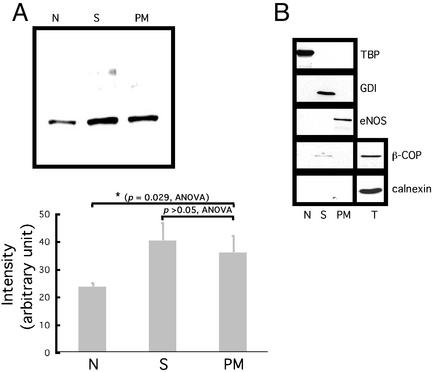
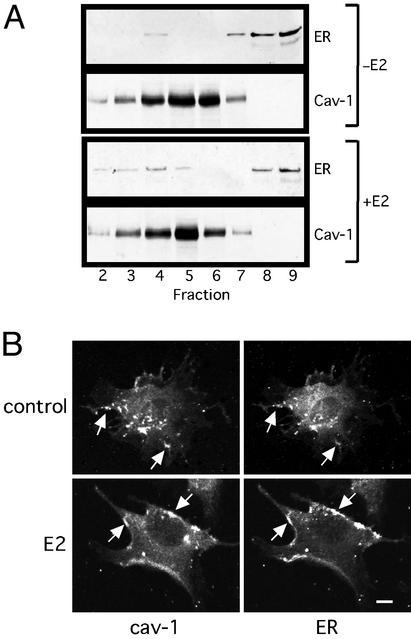
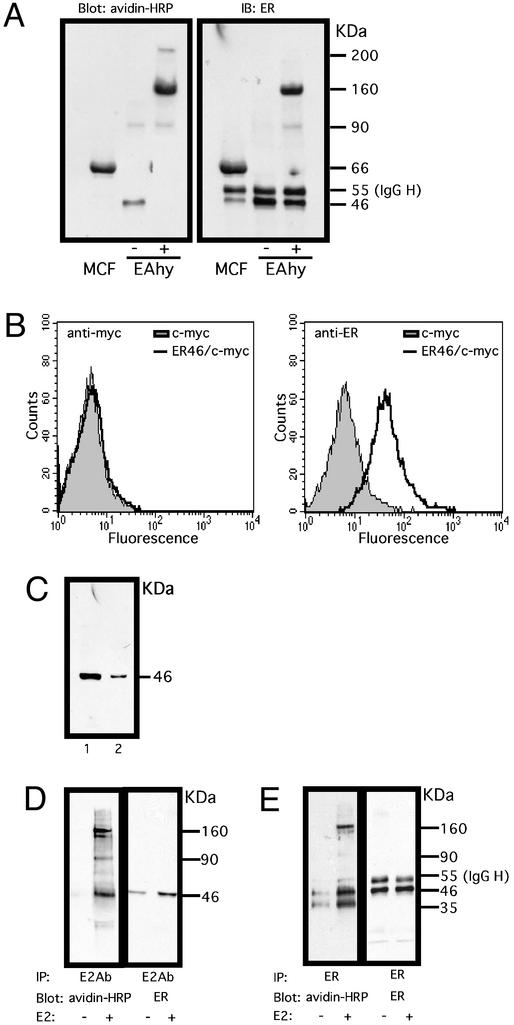
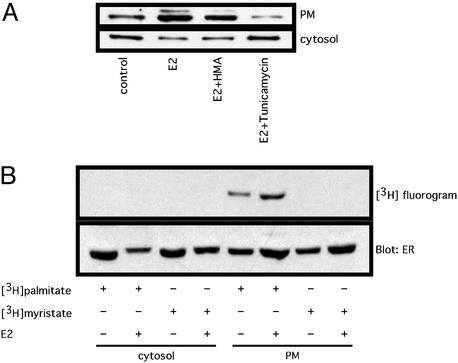
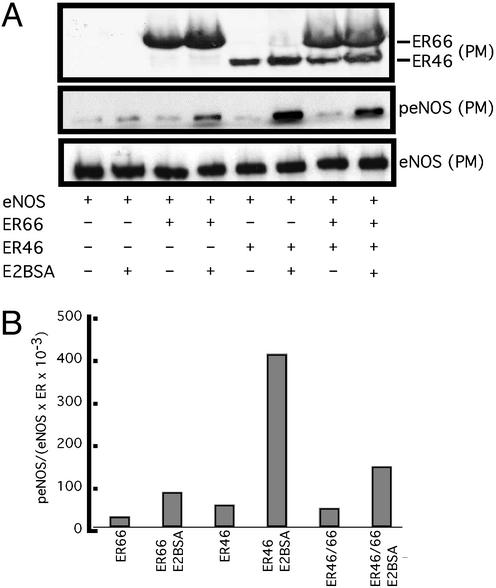
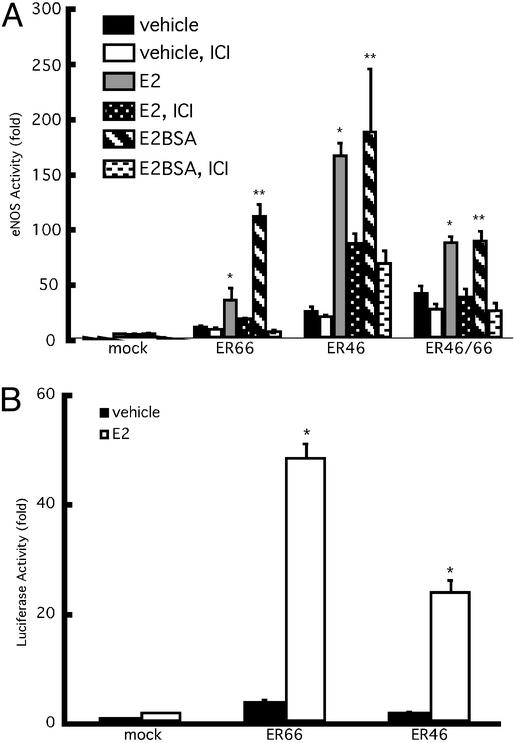
Estrogen receptor (ER) alpha variants have been identified in an array of nonendothelial cells. We previously demonstrated that estrogen rapidly induces nitric oxide release via a phosphatidylinositol 3-kinase/Akt/endothelial nitric-oxide synthase (eNOS) pathway in EA.hy926 cells (immortalized human endothelial cells), which express a 46-kDa ER. We now confirm that, due to alternative splicing, the 46-kDa endothelial cell protein (ER46) is an amino-terminal truncated product of full-length ER alpha (ER66). ER46 is expressed in the plasma membrane, cytosol, and nucleus of resting, estrogen-deprived cells. Flow cytometric and immunofluorescence microscopic analyses demonstrated that the ER46 C but not N terminus is Ab-accessible in the plasma membrane. Inhibition of palmitoylation with tunicamycin and [(3)H]palmitic acid labeling demonstrated an estrogen-induced, palmitoylation-dependent plasma membrane ER46 recruitment, with reorganization into caveolae. In reconstituted, estrogen-stimulated COS-7 (ER-null) cells, membrane ER46 more efficiently triggered membrane eNOS phosphorylation than ER66. Conversely, ER66 more efficiently mediated estrogen response element reporter-gene transactivation than ER46. These results demonstrate that ER46 is localized and further dynamically targeted to the plasma membrane in a palmitoylation-dependent manner. ER46 more efficiently modulates membrane-initiated estrogen actions, including eNOS activation, than full-length ER66. These findings may have important implications in vascular-specific targeting of estrogen receptor agonists.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
