

BIR-1, a Caenorhabditis elegans homologue of Survivin, regulates transcription and development
- PMID: 12682297
- PMCID: PMC154329
- DOI: 10.1073/pnas.0730770100
BIR-1, a Caenorhabditis elegans homologue of Survivin, regulates transcription and development
Abstract
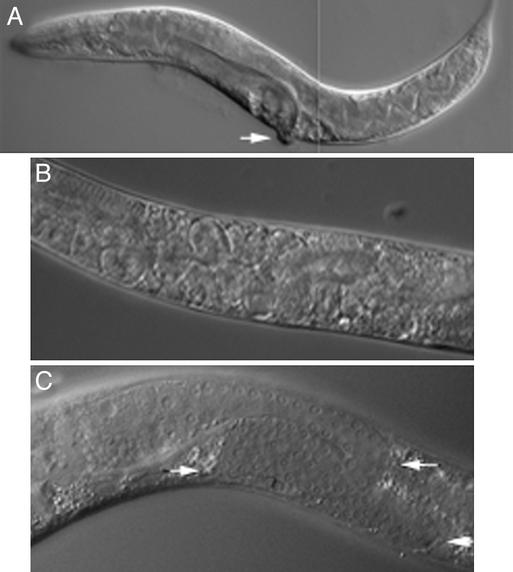
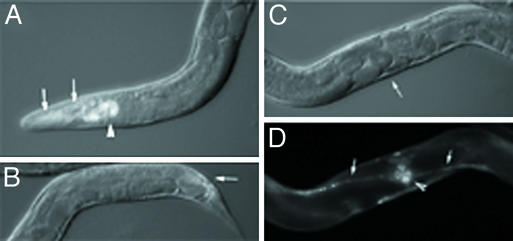
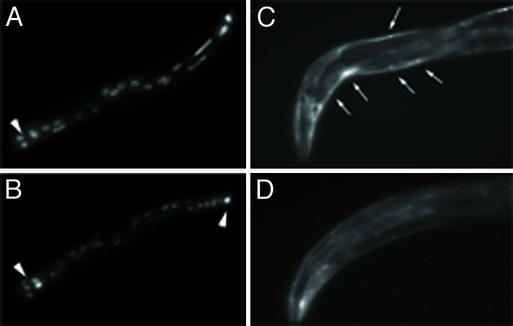
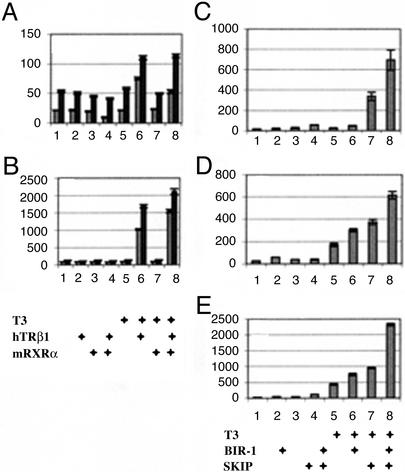
bir-1, a Caenorhabditis elegans inhibitor-of-apoptosis gene homologous to Survivin is organized in an operon with the transcription cofactor C. elegans SKIP (skp-1). Because genes arranged in operons are frequently linked functionally, we have asked whether BIR-1 also functions in transcription. bir-1 inhibition resulted in multiple developmental defects that overlapped with C. elegans SKIP loss-of-function phenotypes: retention of eggs, dumpy, movement defects, and lethality. bir-1 RNA-mediated interference decreased expression of several gfp transgenes and the endogenous genes dpy-7 and hlh-1. Immunoblot analysis revealed decreased phosphoacetylated histones in bir-1 RNA-mediated interference-treated worms. In a heterologous transfection system, BIR-1 augments thyroid hormone-regulated transcription and has an additive effect with SKIP. These results show that BIR-1 functions in the regulation of transcription and development.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
