

Cloning and characterization of tRNA (m1A58) methyltransferase (TrmI) from Thermus thermophilus HB27, a protein required for cell growth at extreme temperatures
- PMID: 12682365
- PMCID: PMC153742
- DOI: 10.1093/nar/gkg314
Cloning and characterization of tRNA (m1A58) methyltransferase (TrmI) from Thermus thermophilus HB27, a protein required for cell growth at extreme temperatures
Abstract
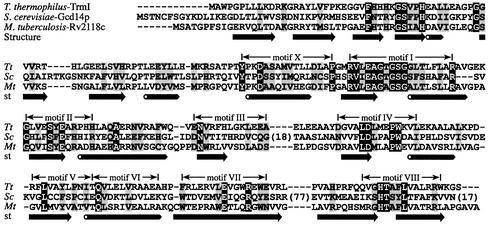
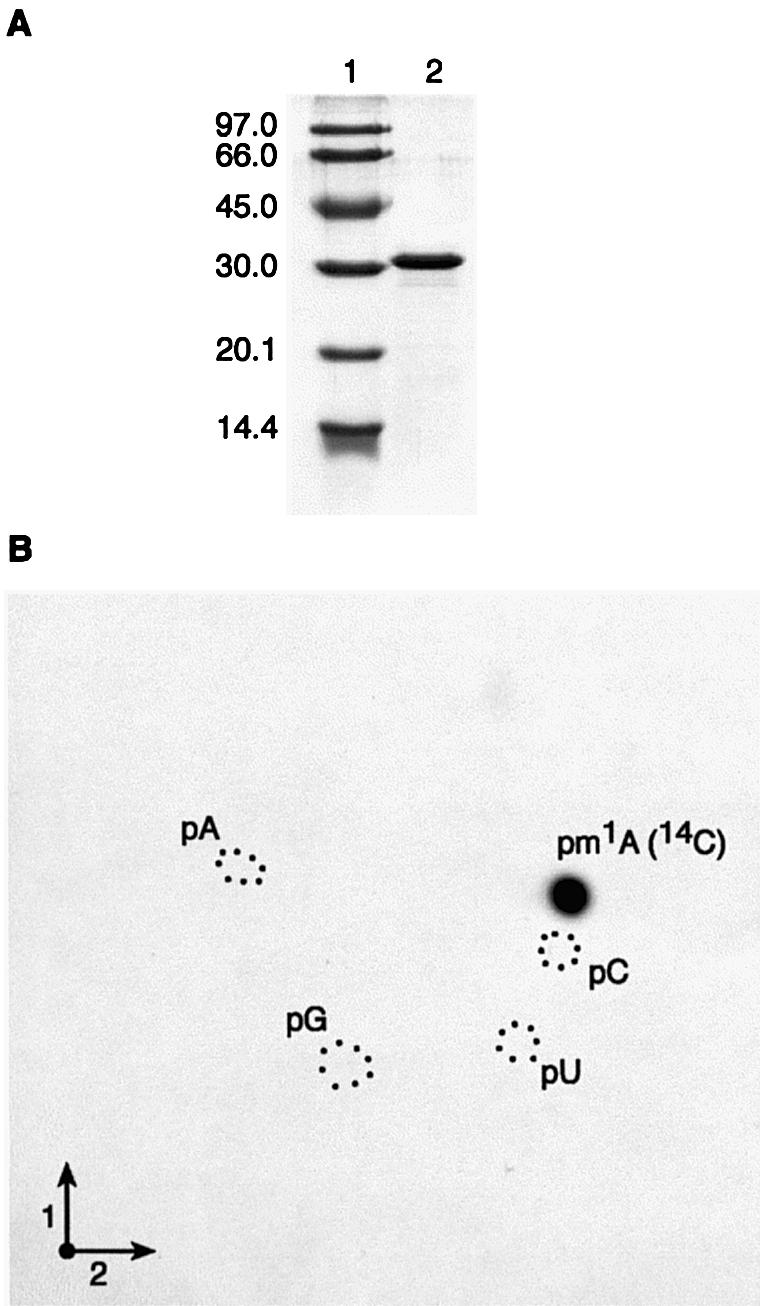
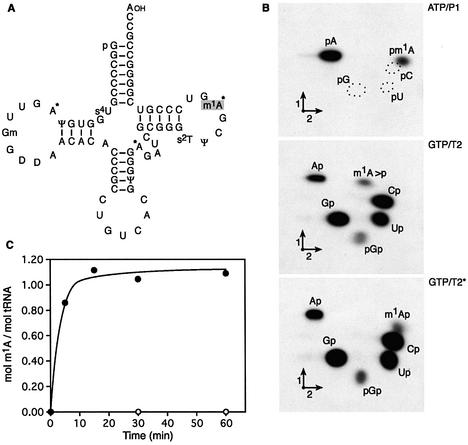
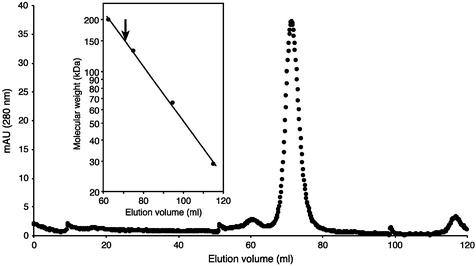
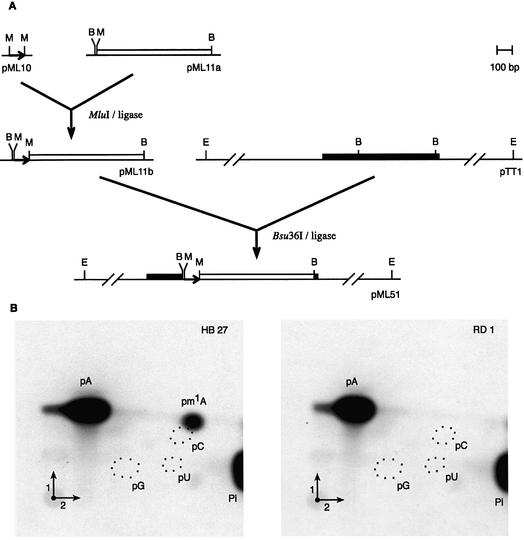
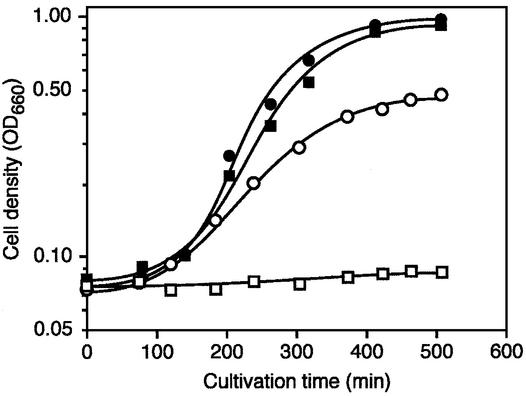
N1-methyladenosine (m1A) is found at position 58 in the T-loop of many tRNAs. In yeast, the formation of this modified nucleoside is catalyzed by the essential tRNA (m1A58) methyltransferase, a tetrameric enzyme that is composed of two types of subunits (Gcd14p and Gcd10p). In this report we describe the cloning, expression and characterization of a Gcd14p homolog from the hyperthermophilic bacterium Thermus thermophilus. The purified recombinant enzyme behaves as a homotetramer of 150 kDa by gel filtration and catalyzes the site- specific formation of m1A at position 58 of the T-loop of tRNA in the absence of any other complementary protein. S-adenosylmethionine is used as donor of the methyl group. Thus, we propose to name the bacterial enzyme TrmI and accordingly its structural gene trmI. These results provide a key evolutionary link between the functionally characterized two-component eukaryotic enzyme and the recently described crystal structure of an uncharacterized, putative homotetrameric methyltransferase Rv2118c from Mycobacterium tuberculosis. Interest ingly, inactivation of the T.thermophilus trmI gene results in a thermosensitive phenotype (growth defect at 80 degrees C), which suggests a role of the N1-methylation of tRNA adenosine-58 in adaptation of life to extreme temperatures.
Figures
References
-
- Baguley B.C. and Staehelin,M. (1968) Substrate specificity of adenine-specific transfer RNA methylase in normal and leukemic tissues. Eur. J. Biochem., 6, 1–7. - PubMed
-
- Kuchino Y. and Nishimura,S. (1974) Methylation of Escherichia coli transfer ribonucleic acids by adenylate residue-specific transfer ribonucleic acid methylase from rat liver. Biochemistry, 13, 3683–3688. - PubMed
-
- Glick J.M. and Leboy,P.S. (1977) Purification and properties of tRNA (adenine-1)-methyltransferase from rat liver. J. Biol. Chem., 252, 4790–4795. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
