

A cell surface receptor complex for fibrillar beta-amyloid mediates microglial activation
- PMID: 12684452
- PMCID: PMC6742111
- DOI: 10.1523/JNEUROSCI.23-07-02665.2003
A cell surface receptor complex for fibrillar beta-amyloid mediates microglial activation
Abstract
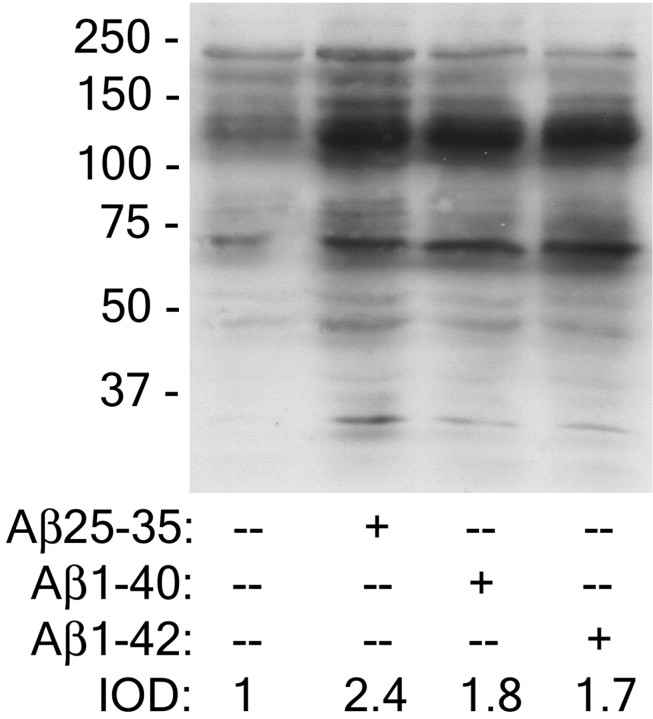
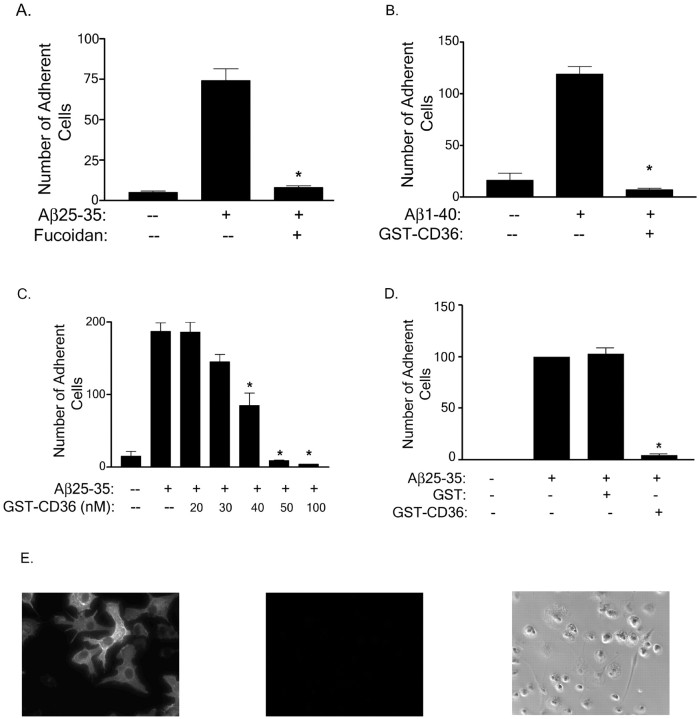
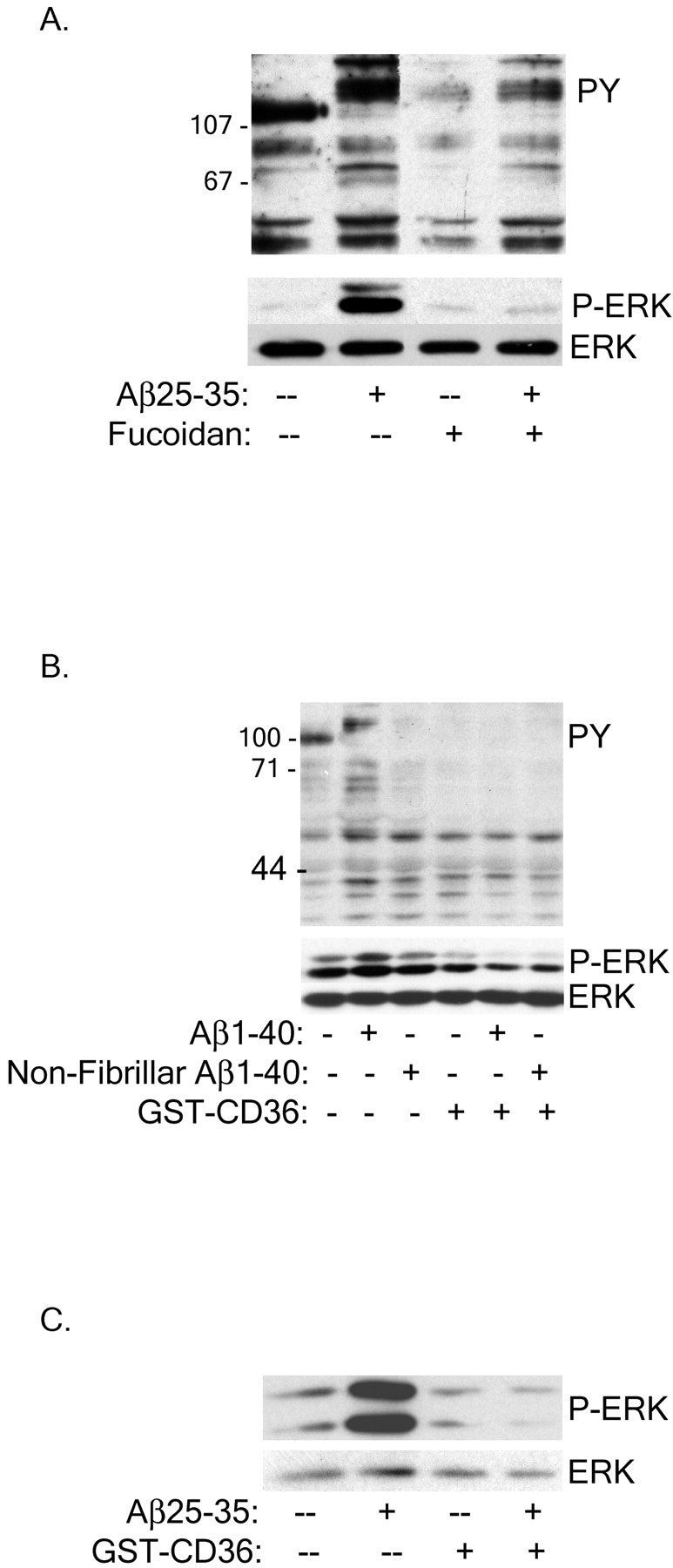
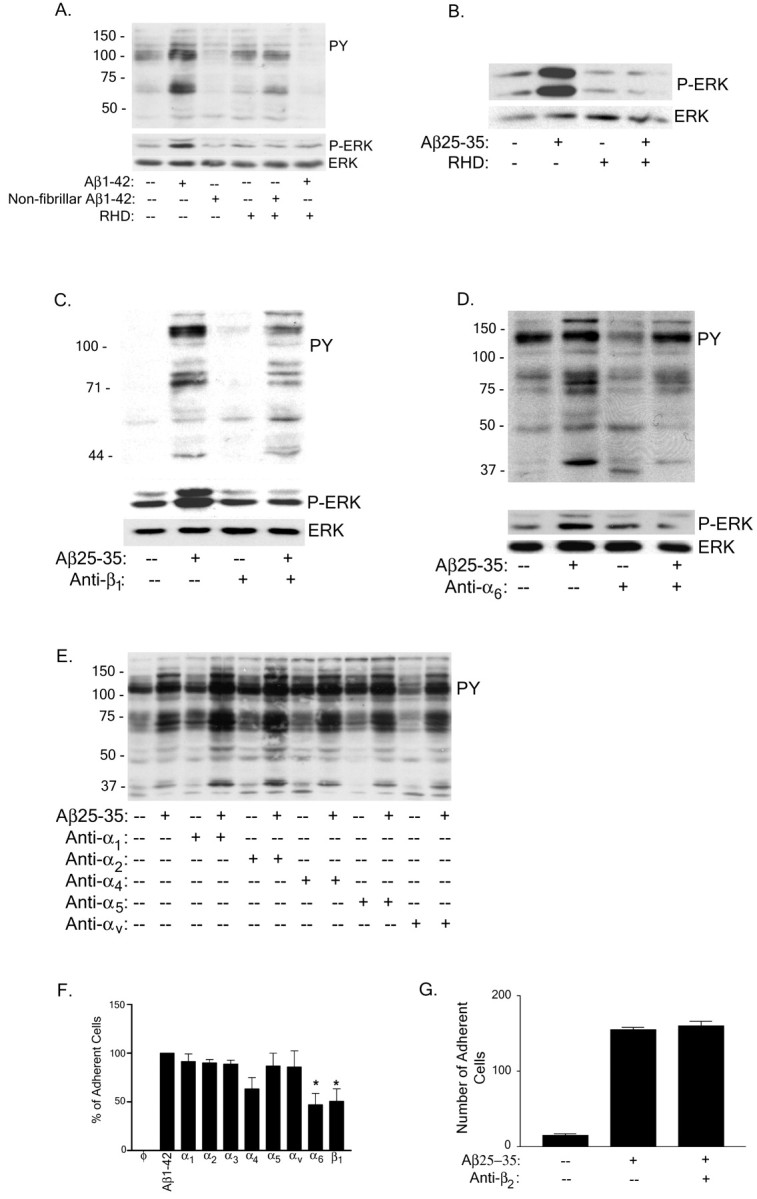
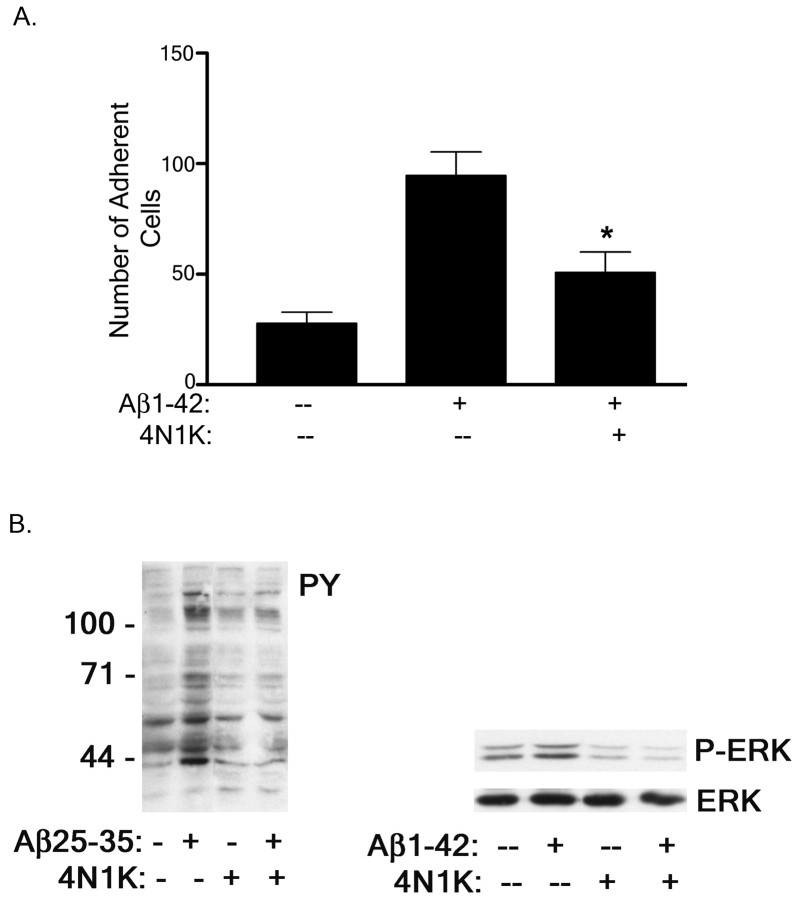
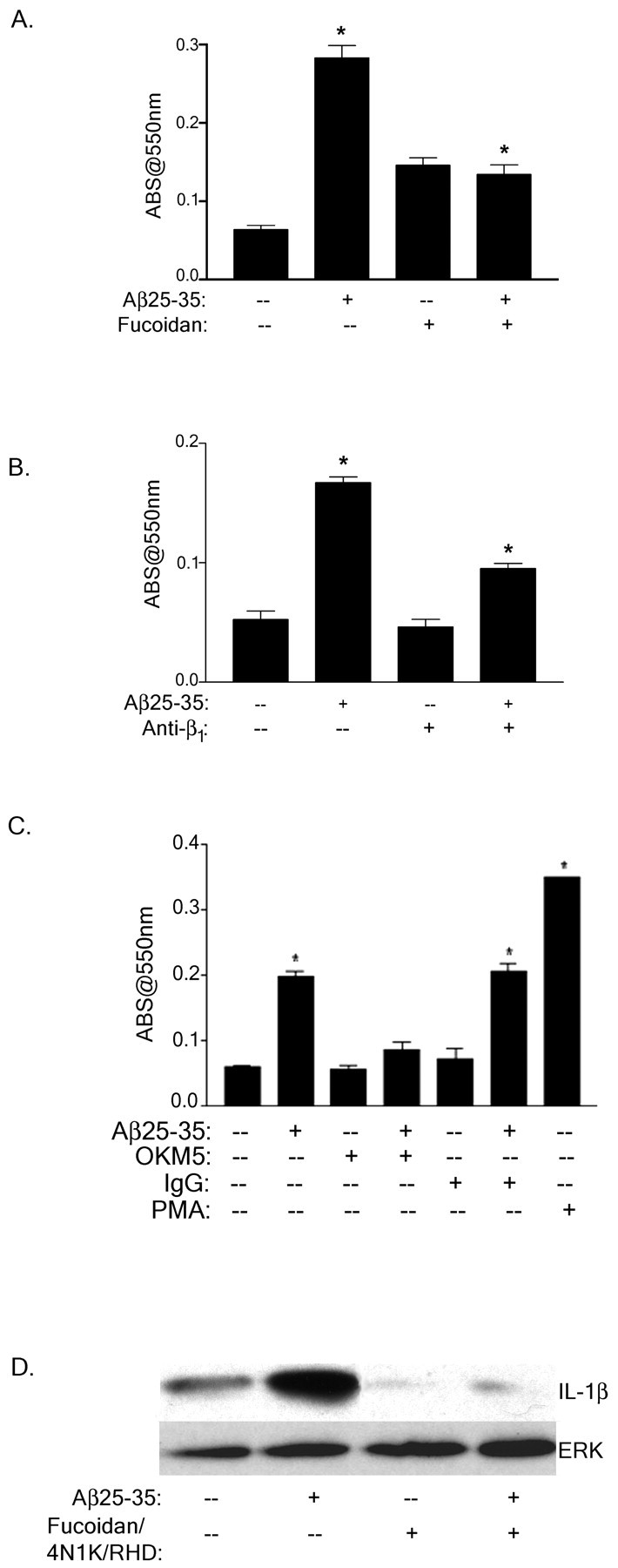
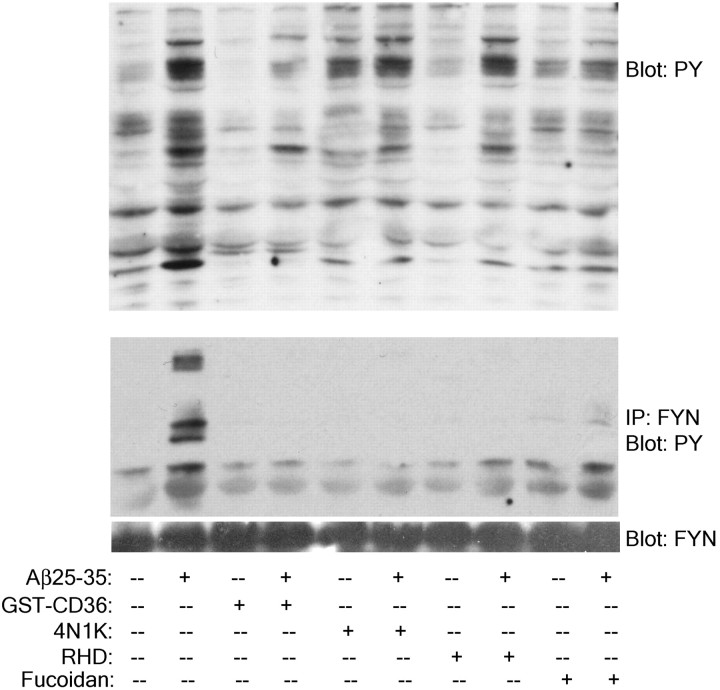
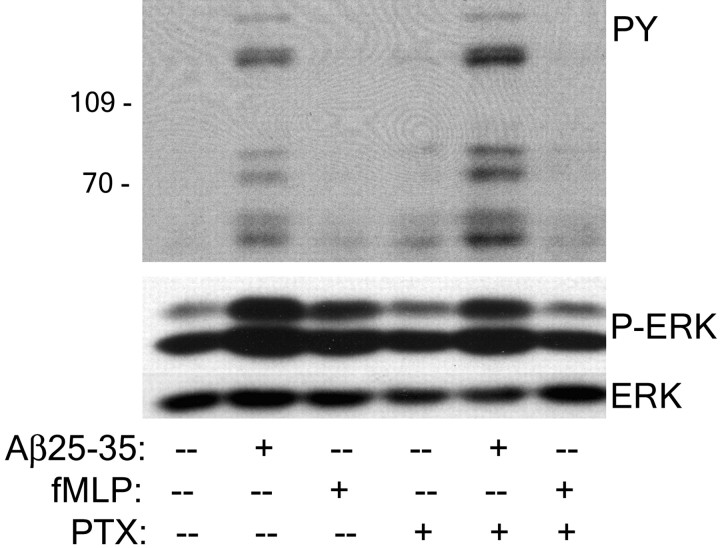
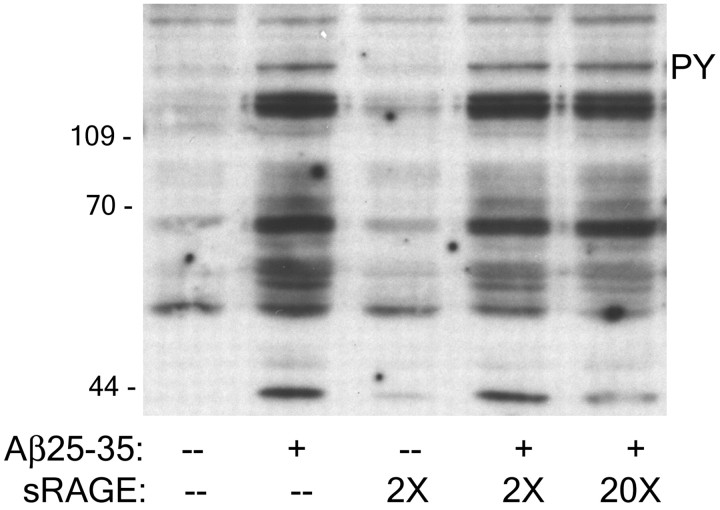
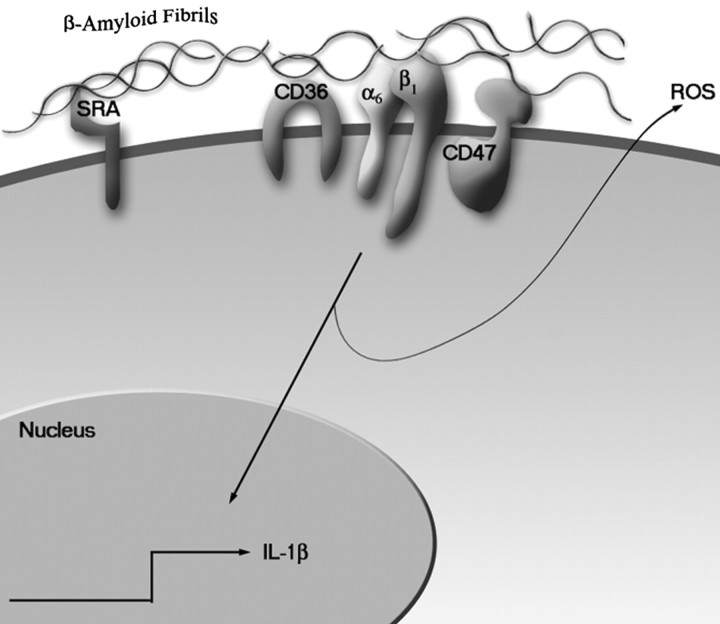
Senile plaques found in the Alzheimer's disease brain are foci of local inflammatory reactions mediated by plaque-associated microglia. The interaction of microglia with compacted deposits of beta-amyloid (Abeta) fibrils results in the stimulation of intracellular Tyr kinase-based signaling cascades and cellular activation, leading to the secretion of proinflammatory molecules. This study identifies a cell surface receptor complex that mediates the binding of microglia to Abeta fibrils and the subsequent activation of intracellular signaling pathways leading to a proinflammatory response. The receptor complex includes the B-class scavenger receptor CD36, the integrin-associated protein/CD47, and the alpha(6)beta(1)-integrin. Antagonists of scavenger receptors, CD36, CD47, and alpha(6)beta(1) inhibited the adhesion of THP-1 monocytes to Abeta fibrils. In addition, peptide competitors of Abeta fibril interactions with CD36, scavenger receptors, CD47, and the alpha(6)beta(1)-integrin inhibited Abeta stimulation of Tyr kinase-based signaling cascades in both THP-1 monocytes and murine microglia as well as interleukin 1beta production. A scavenger receptor antagonist and antibodies specific for CD36 and the beta(1)-integrin subunit also inhibited the Abeta-stimulated generation of reactive oxygen species. Importantly, the principal components of this receptor complex are shared with those for other fibrillar proteins and thus represent general elements through which myeloid lineage cells recognize complex fibrillar proteins. Identification of the cell surface molecules that interact with Abeta fibrils and mediate their activation of intracellular signaling cascades represents a potential intervention point in the treatment of Alzheimer's disease.
Figures
References
-
- Akiyama H, Barger S, Barnum S, Bradt B, Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich BL, Finch CE, Frautschy S, Griffin WS, Hampel H, Hull M, Landreth G, Lue L, Mrak R, Mackenzie IR, McGeer PL, O'Banion MK, Pachter J, Pasinetti G, Plata-Salaman C, Rogers J, Rydel R, Shen Y, Streit W, Strohmeyer R, Tooyoma I, Van Muiswinkel FL, Veerhuis R, Walker D, Webster S, Wegrzyniak B, Wenk G, Wyss-Coray T. Inflammation and Alzheimer's disease. Neurobiol Aging. 2000;21:383–421. - PMC - PubMed
-
- Araujo DM, Cotman CW. Beta-amyloid stimulates glial cells in vitro to produce growth factors that accumulate in senile plaques in Alzheimer's disease. Brain Res. 1992;569:141–145. - PubMed
-
- Babic I, Schallhorn A, Lindberg FP, Jirik FR. SHPS-1 induces aggregation of Ba/F3 pro-B cells via an interaction with CD47. J Immunol. 2000;164:3652–3658. - PubMed
-
- Bianca VD, Dusi S, Bianchini E, Dal Pra I, Rossi F. beta-amyloid activates the O-2 forming NADPH oxidase in microglia, monocytes, and neutrophils: a possible inflammatory mechanism of neuronal damage in Alzheimer's disease. J Biol Chem. 1999;274:15493–15499. - PubMed
-
- Boland K, Behrens M, Choi D, Manias K, Perlmutter DH. The serpin-enzyme complex receptor recognizes soluble, nontoxic amyloid-β peptide but not aggregated, cytotoxic amyloid-β peptide. J Biol Chem. 1996;271:18032–18044. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials