

Dopamine D1-class receptors selectively modulate a slowly inactivating potassium current in rat medial prefrontal cortex pyramidal neurons
- PMID: 12684454
- PMCID: PMC6742102
- DOI: 10.1523/JNEUROSCI.23-07-02686.2003
Dopamine D1-class receptors selectively modulate a slowly inactivating potassium current in rat medial prefrontal cortex pyramidal neurons
Abstract
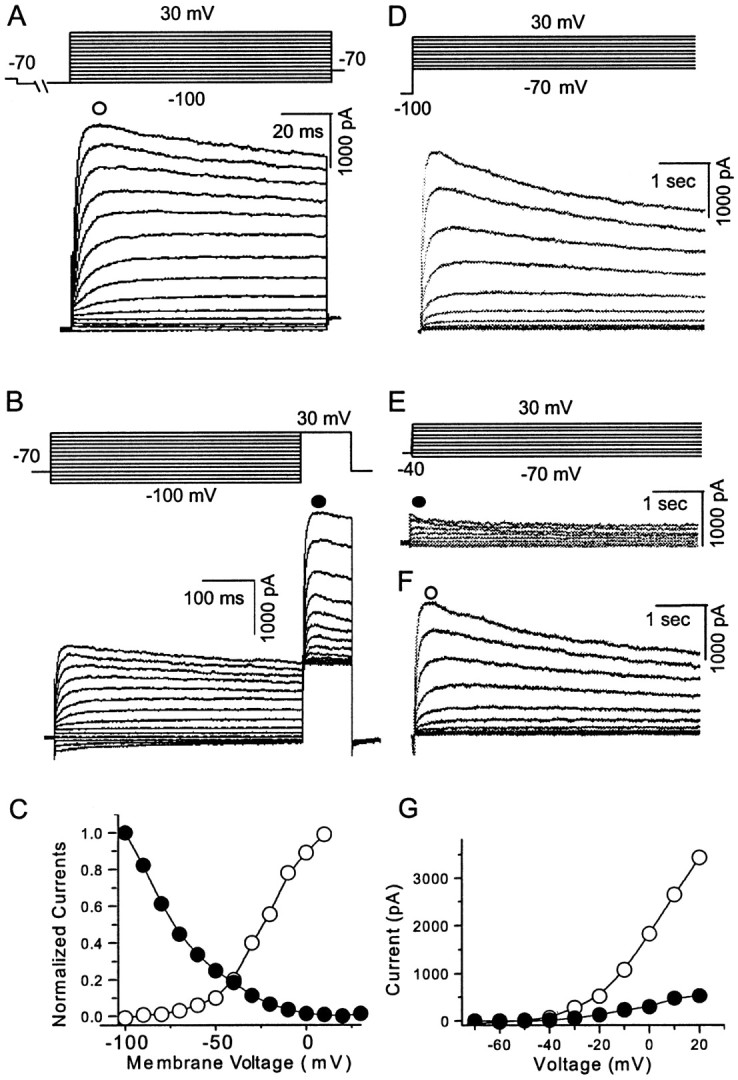
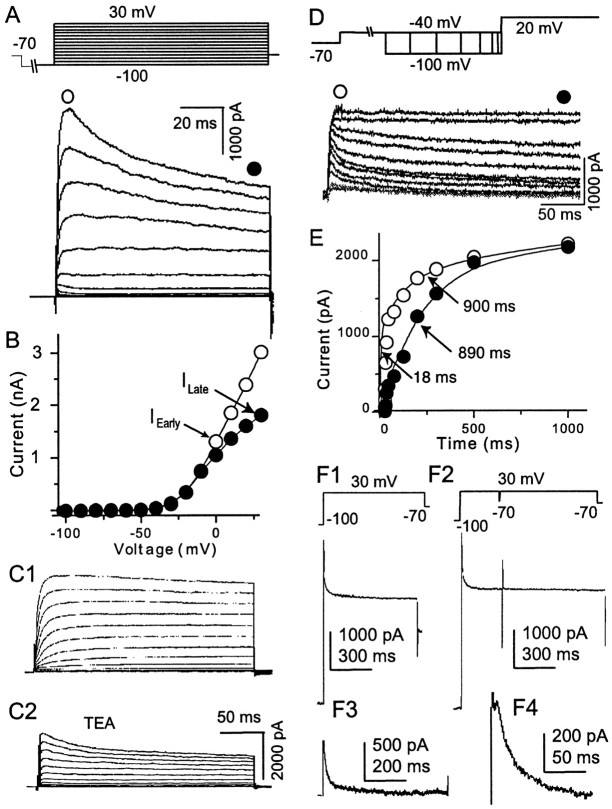
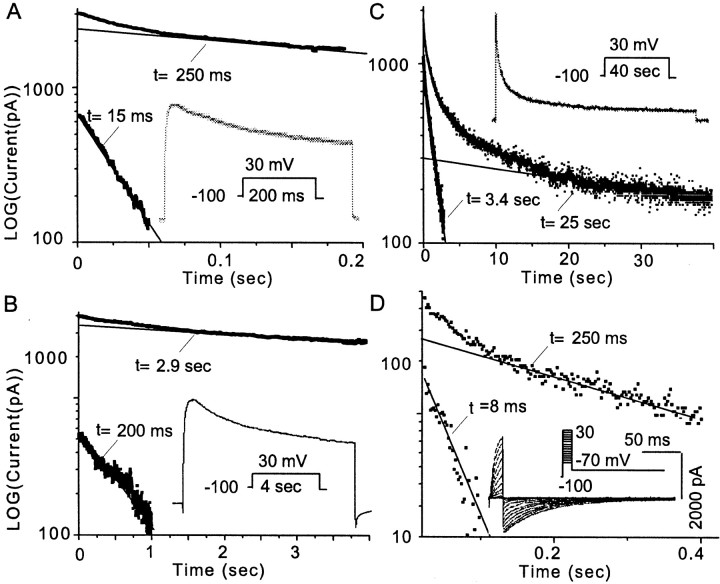
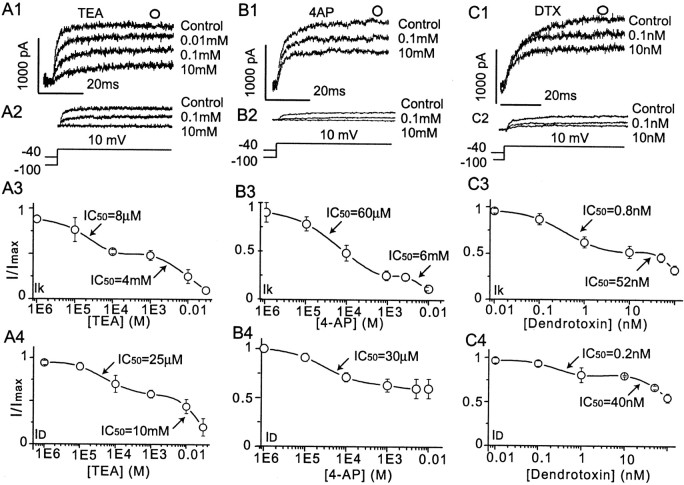
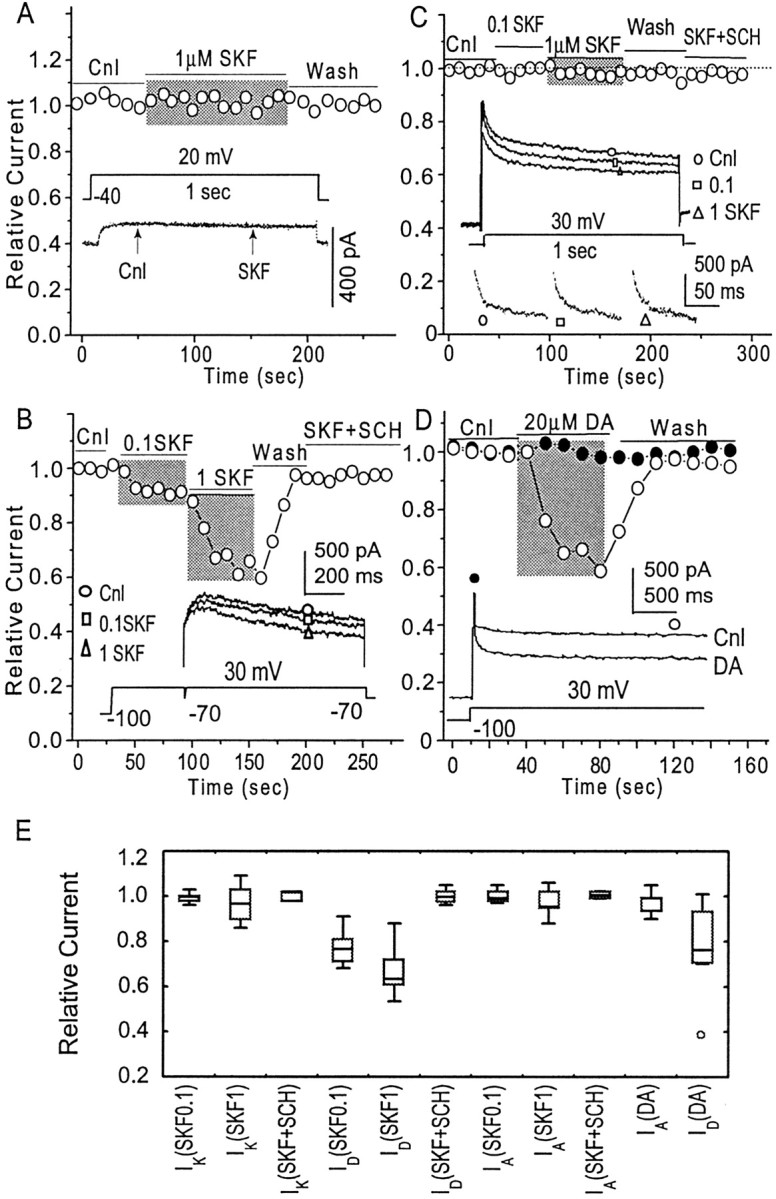
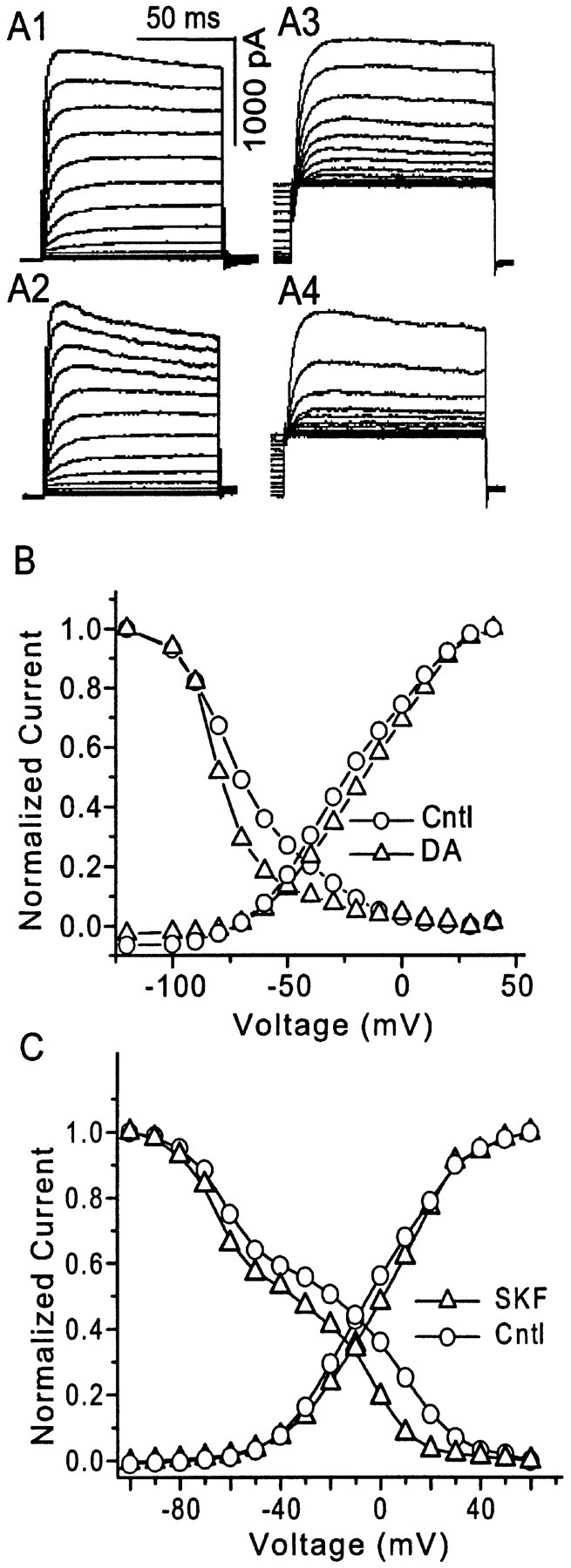
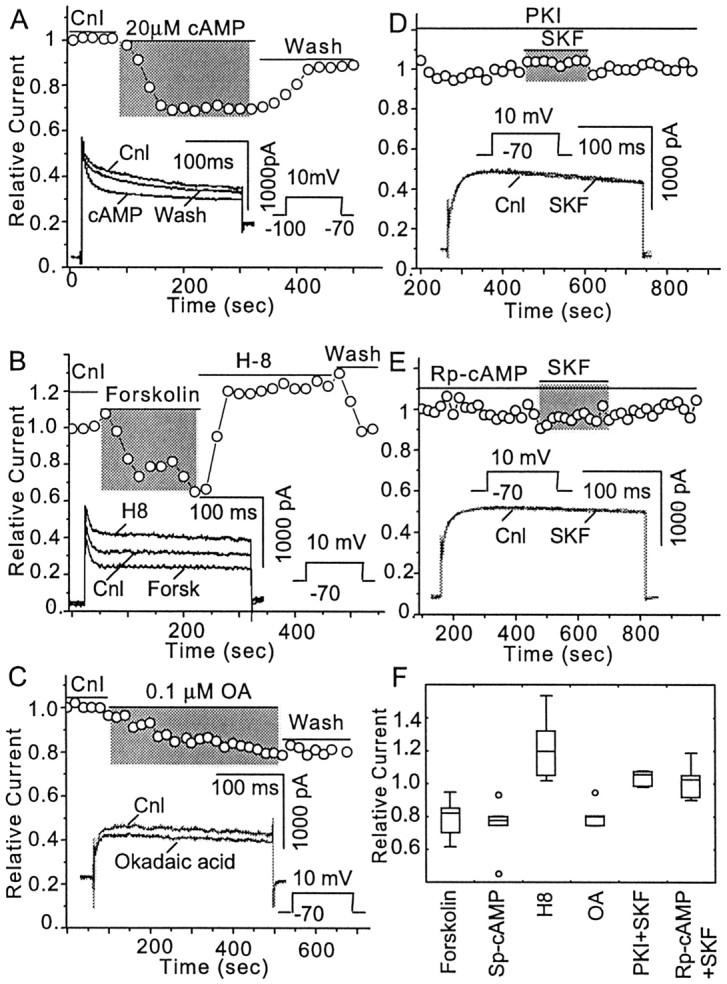
The dopamine (DA) innervation of medial prefrontal cortex (mPFC) regulates cognitive activity in a complex manner. Alterations of DA function, particularly via the DA D1 receptor class (D1R), are implicated in both schizophrenia and drug addiction, yet the precise roles of DA in modulating mPFC excitability remain unclear. We focused on DA modulation of voltage-gated K(+) current (VGKC) in acutely dissociated rat mPFC pyramidal neurons. We defined three components of the whole-cell VGKC according to biophysical and pharmacological properties. The A-type current (I(A)), with rapid activation and inactivation kinetics, was completely inactivated by prolonged holding of the membrane potential at -40 mV and was sensitive to the K(+) channel blocker 4-aminopyridine (4-AP) but not tetraethylammonium (TEA) or dendrotoxin (DTX). The slowly inactivating K(+) current (I(D)), with rapid activation but relatively slow inactivation, was the major contributor to VGKC and was completely inactivated at -40 mV and sensitive to TEA and DTX but less so to 4-AP. The very slowly inactivating K(+) current (I(K)) was elicited by command steps to more depolarized potentials from a prolonged holding potential of -40 mV and was sensitive to all three blockers. Stimulation of DA D2 receptors failed to alter any component of whole-cell VGKC. Stimulation of DA D1Rs selectively suppressed I(D), an effect mimicked by the adenylyl cyclase activator forskolin, the active cAMP analog Sp-cAMP, and the protein phosphatase inhibitor okadaic acid. Inhibition of protein kinase A (PKA) with either PKI or Rp-cAMP abolished D1R modulation. Thus, the DA D1R/cAMP/PKA signaling pathway mediates modulation of I(D) by DA in rat mPFC pyramidal neurons.
Figures
References
-
- Berger B, Gaspar P, Verney C. Dopaminergic innervation of the cerebral cortex: unexpected differences between rodents and primates. Trends Neurosci. 1991;14:21–27. - PubMed
-
- Bjorklund A, Divac I, Lindvall O. Regional distribution of catecholamines in monkey cerebral cortex, evidence for a dopaminergic innervation of the primate prefrontal cortex. Neurosci Lett. 1978;7:115–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources