

Spinal cats on the treadmill: changes in load pathways
- PMID: 12684465
- PMCID: PMC6742096
- DOI: 10.1523/JNEUROSCI.23-07-02789.2003
Spinal cats on the treadmill: changes in load pathways
Abstract
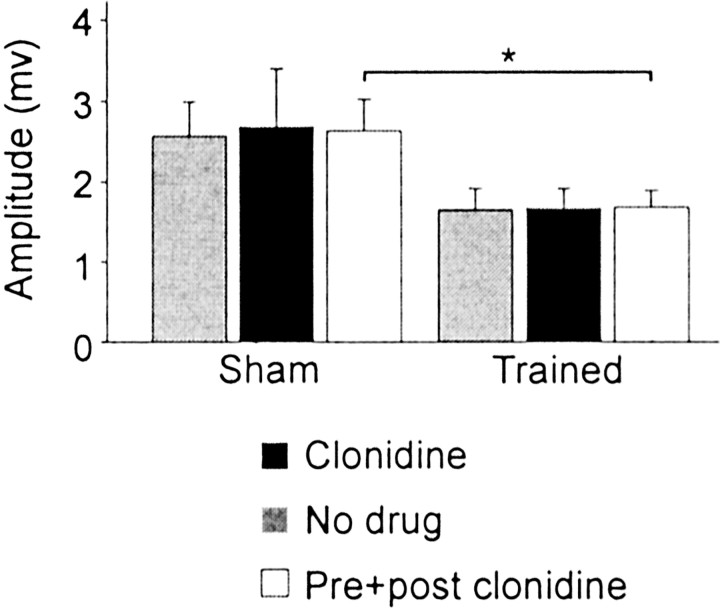
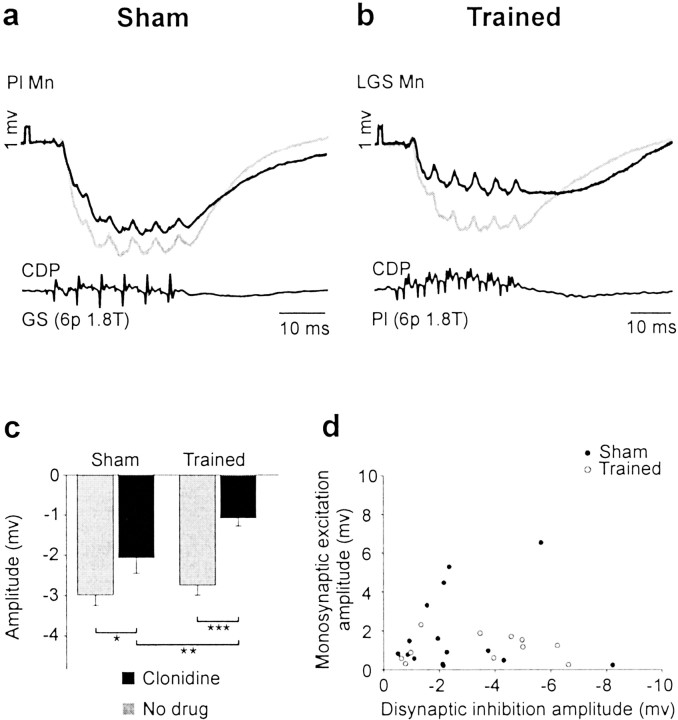
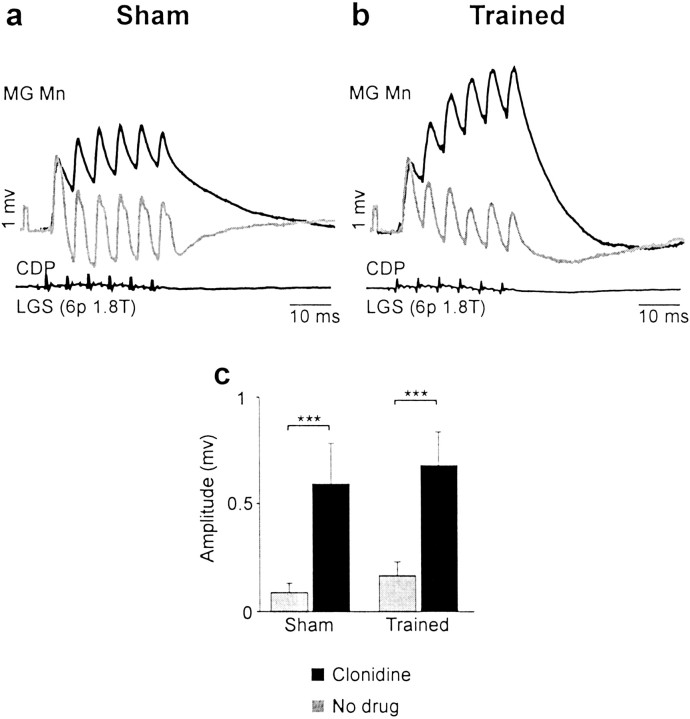
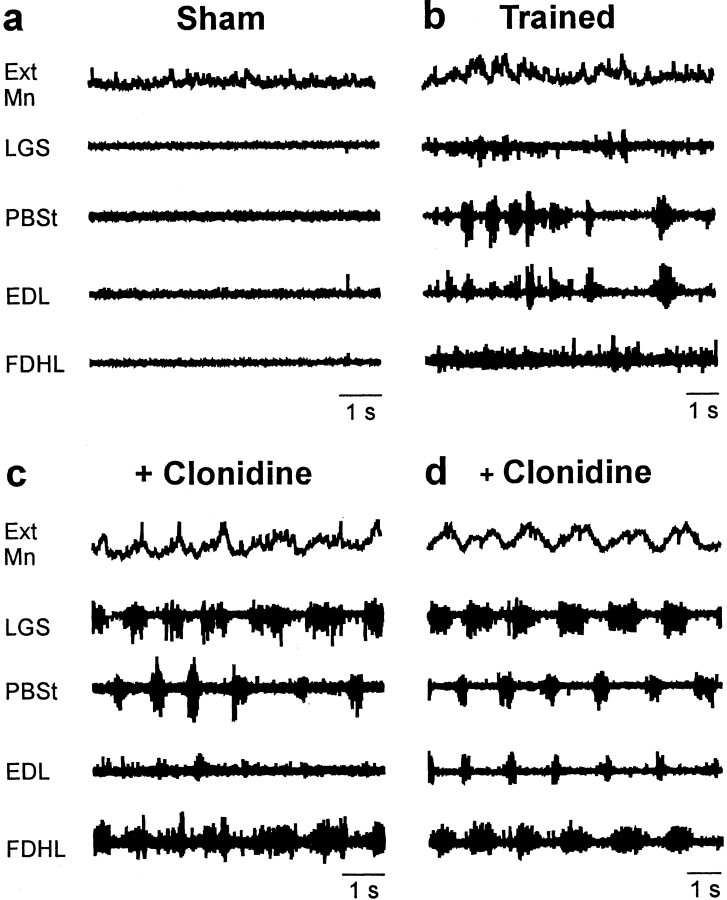
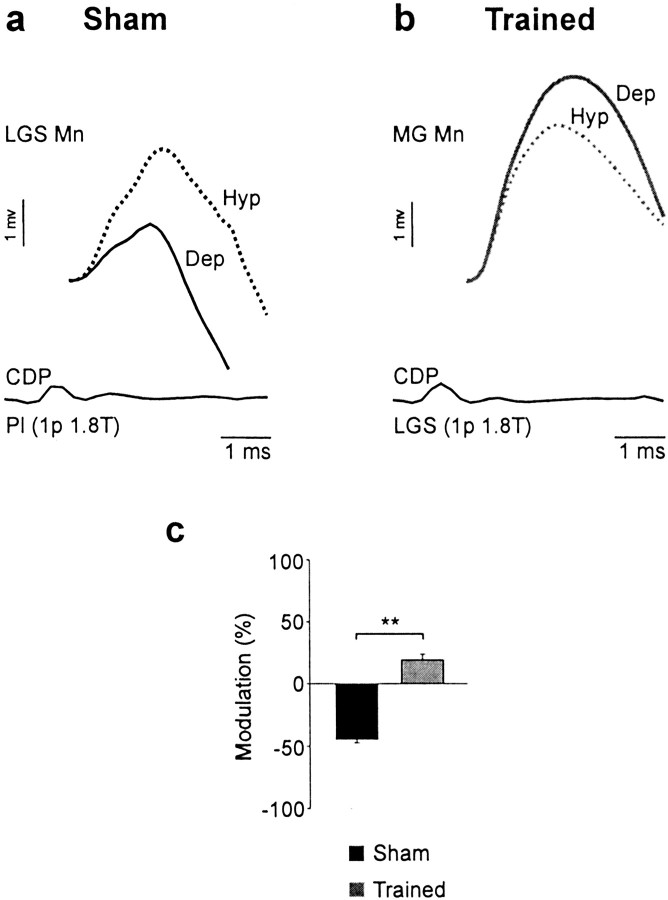
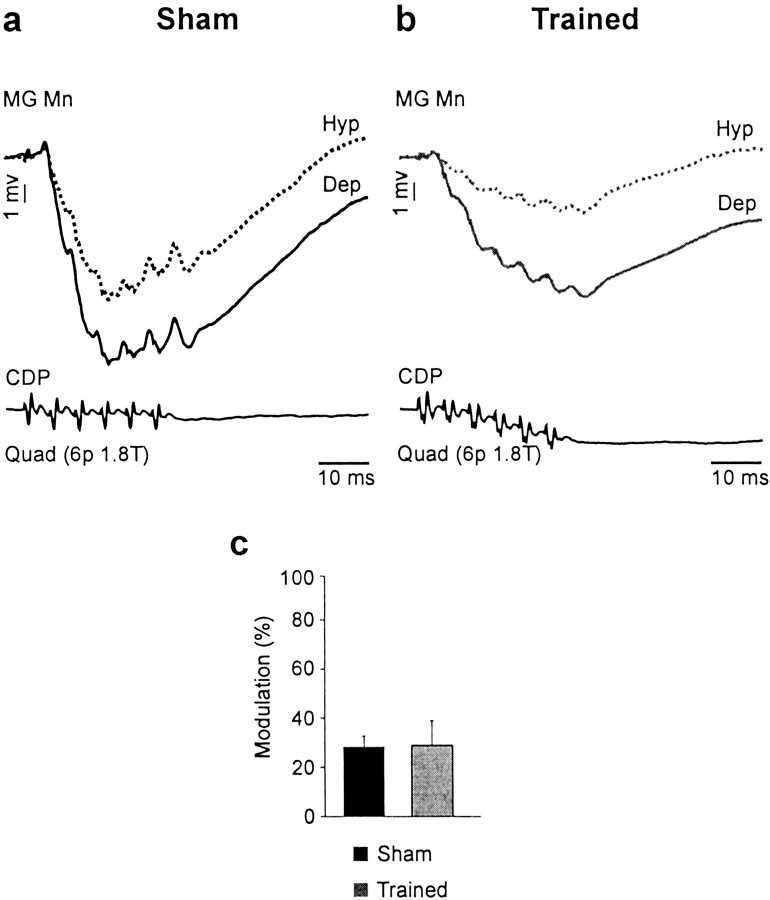
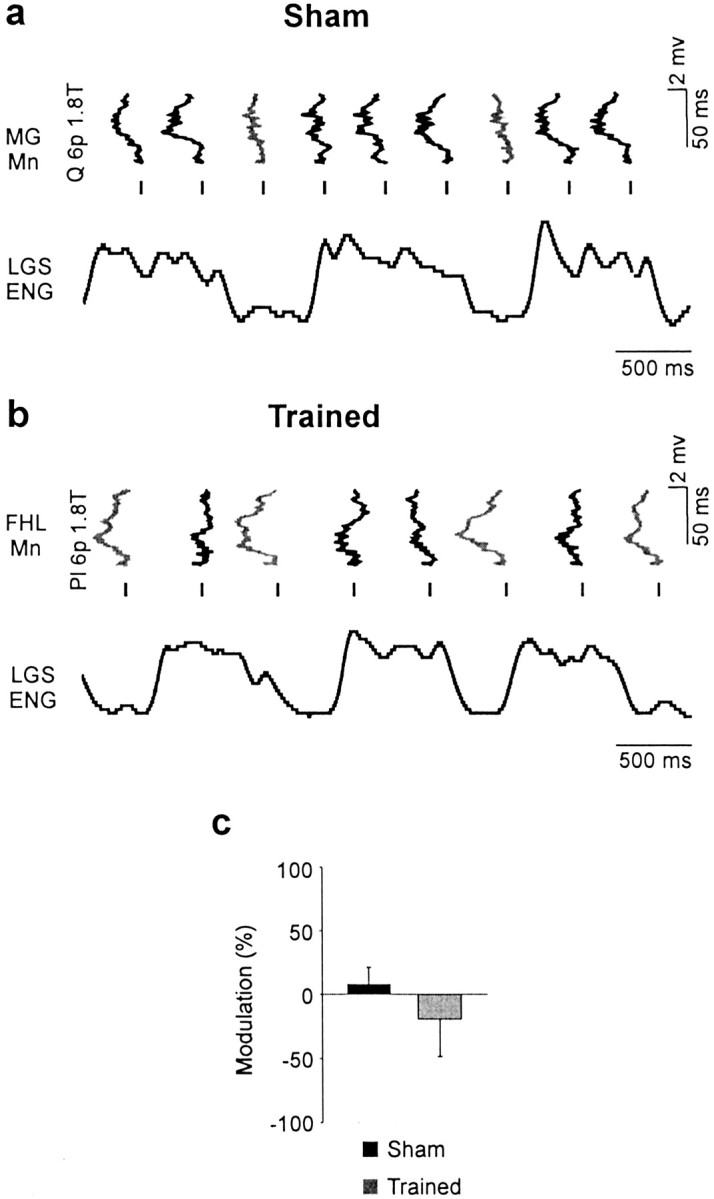
Treadmill training and clonidine, an alpha-2 noradrenergic agonist, have been shown to improve locomotion after spinal cord injury. We speculate that transmission in load pathways, which are involved in body support during stance, is specifically modified by training. This was evaluated by comparing two groups of spinal cats; one group (n = 11) was trained to walk until full-weight-bearing (3-4 weeks), and the other (shams; n = 7) was not. During an acute experiment, changes in group I pathways, monosynaptic excitation, disynaptic inhibition, and polysynaptic excitation were investigated by measuring the response amplitude in extensor motoneurons before and after clonidine injection. Monosynaptic excitation was not modified by clonidine but was decreased significantly by training. Disynaptic inhibition was significantly decreased by clonidine in both groups, but more significantly in trained cats, and significantly reduced by training after clonidine. Also, clonidine could reverse group IB inhibition into polysynaptic excitation in both groups but more frequently in trained cats. We also investigated whether fictive stepping revealed additional changes. In trained cats, the phase-dependent modulation of all three responses was similar to patterns reported previously, but in shams, modulation of monosynaptic and polysynaptic responses was not. Overall, training appears to decrease monosynaptic excitation and enhance the effects of clonidine in the reduction of disynaptic inhibition and reversal to polysynaptic excitation. Because it is believed that polysynaptic excitatory group I pathways transmit locomotor drive to extensor motoneurons, we suggest that the latter changes would facilitate the recruitment of extensor muscles for recovering weight-bearing during stepping.
Figures

References
-
- Baldissera F, Hultborn H, Illert M. Integration in spinal neuronal systems. In: Brookhart JM, Mountcastle VB, editors. Handbook of physiology. The nervous system, Vol II. American Physiological Society; Bethesda, MD: 1981. pp. 509–595.
-
- Barbeau H, Rossignol S. Recovery of locomotion after chronic spinalization in the adult cat. Brain Res. 1987;412:84–95. - PubMed
-
- Barbeau H, Rossignol S. Initiation and modulation of the locomotor pattern in the adult chronic spinal cat by noradrenergic, serotonergic and dopaminergic drugs. Brain Res. 1991;546:250–260. - PubMed
-
- Barbeau H, Julien C, Rossignol S. The effects of clonidine and yohimbine on locomotion and cutaneous reflexes in the adult chronic spinal cat. Brain Res. 1987;437:83–96. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous