

A chondroitin sulfate proteoglycan PTPzeta /RPTPbeta regulates the morphogenesis of Purkinje cell dendrites in the developing cerebellum
- PMID: 12684467
- PMCID: PMC6742081
- DOI: 10.1523/JNEUROSCI.23-07-02804.2003
A chondroitin sulfate proteoglycan PTPzeta /RPTPbeta regulates the morphogenesis of Purkinje cell dendrites in the developing cerebellum
Abstract
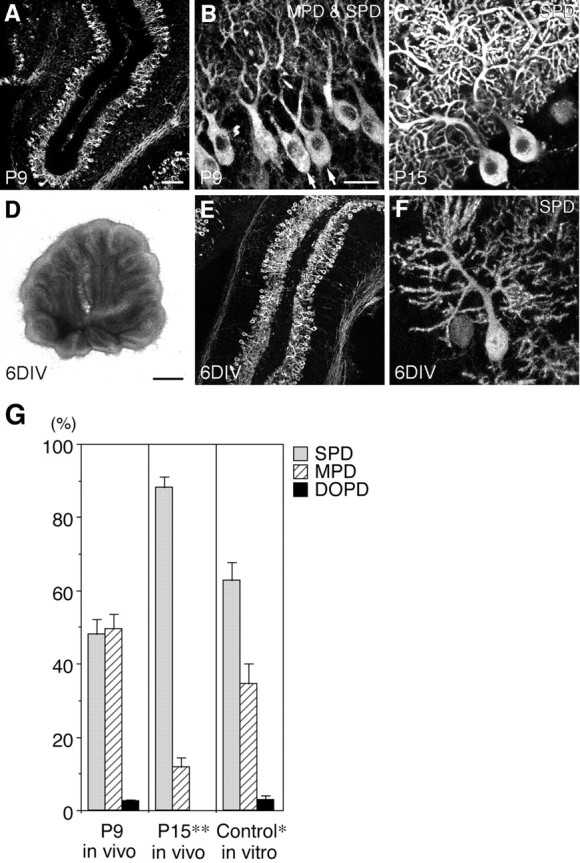
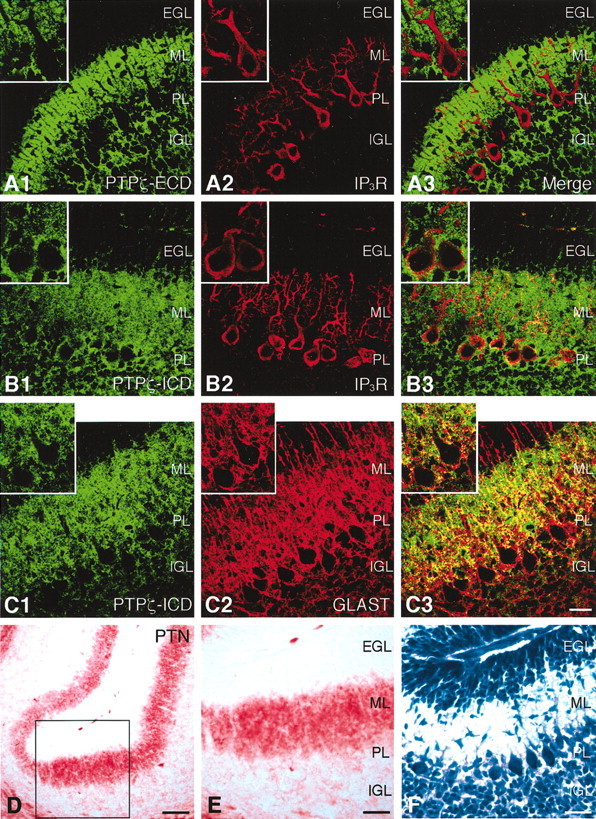
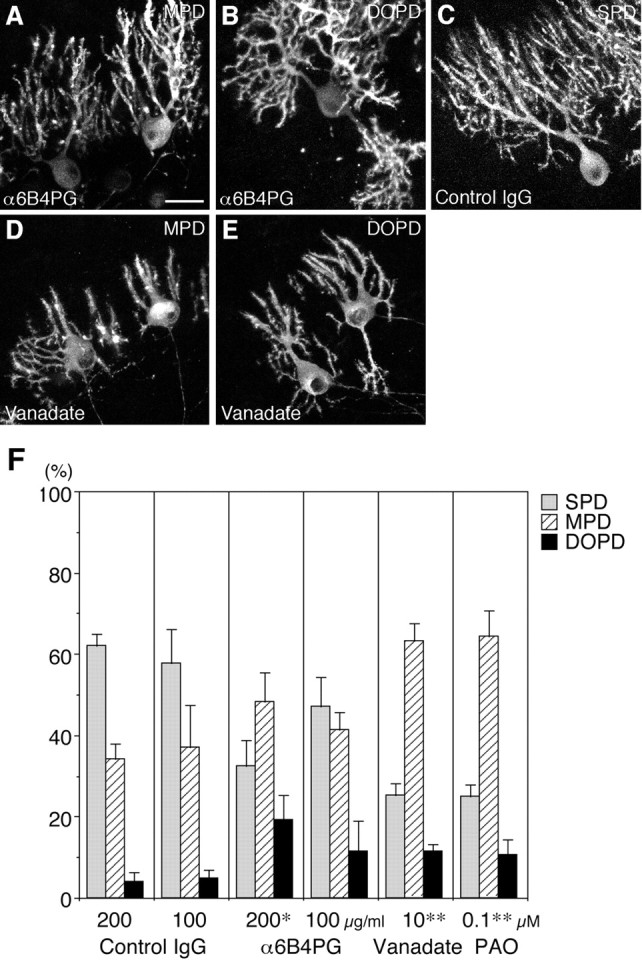
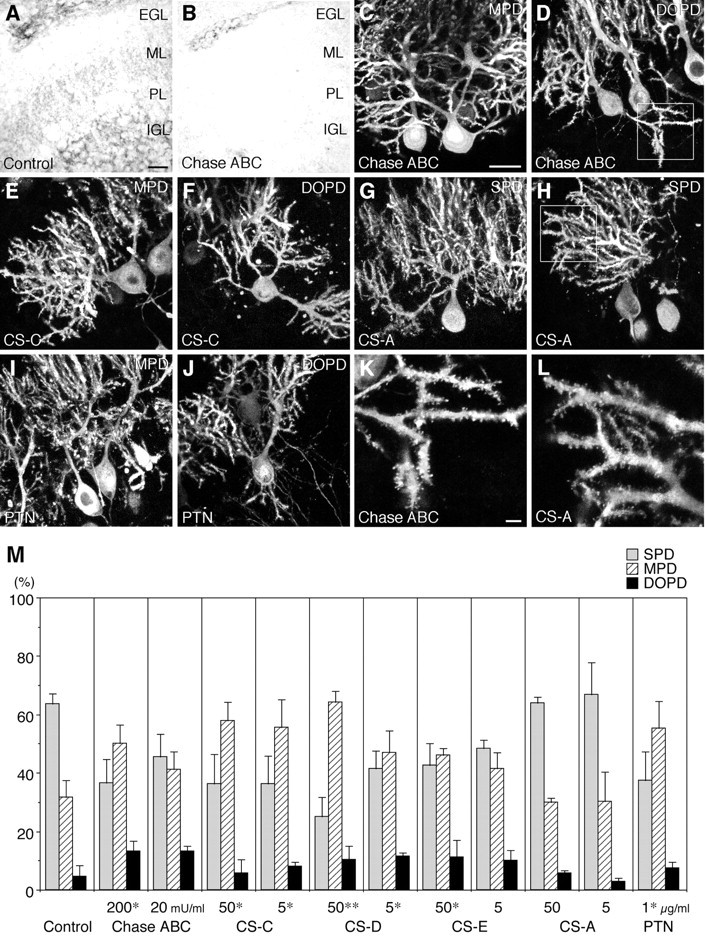
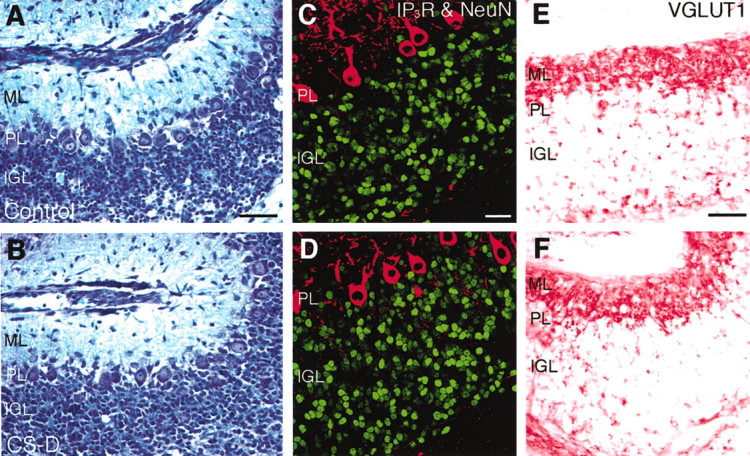
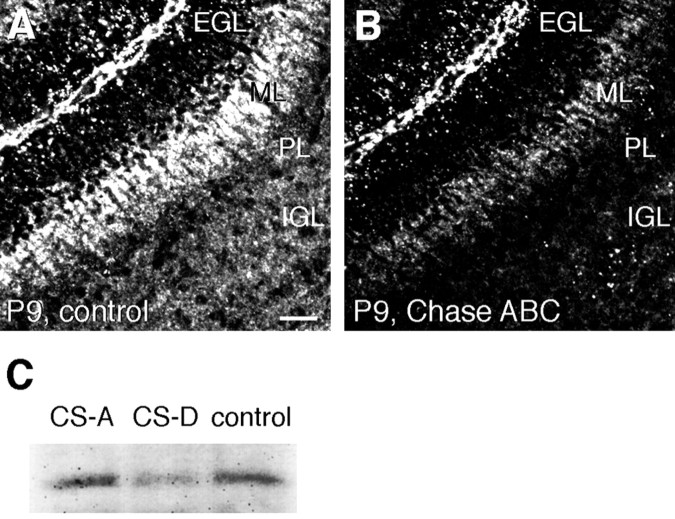
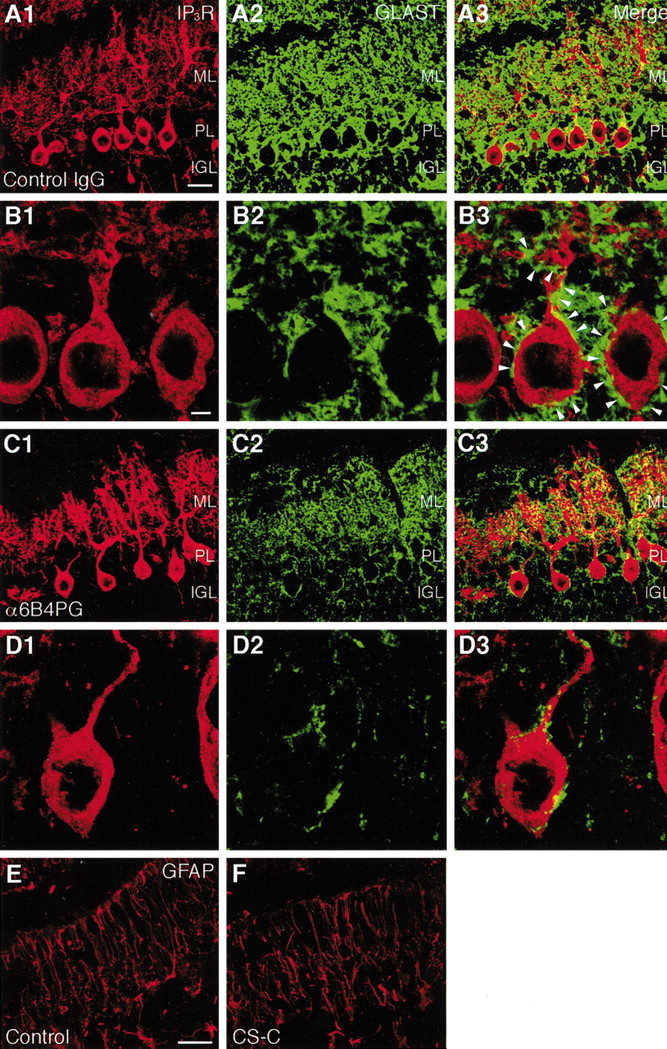
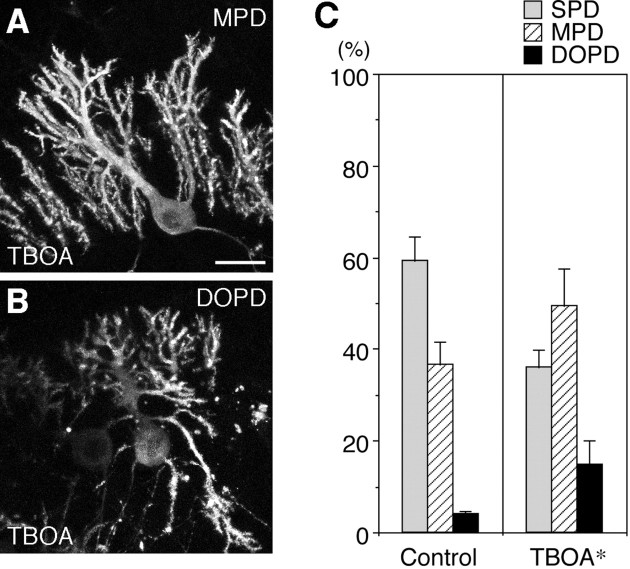
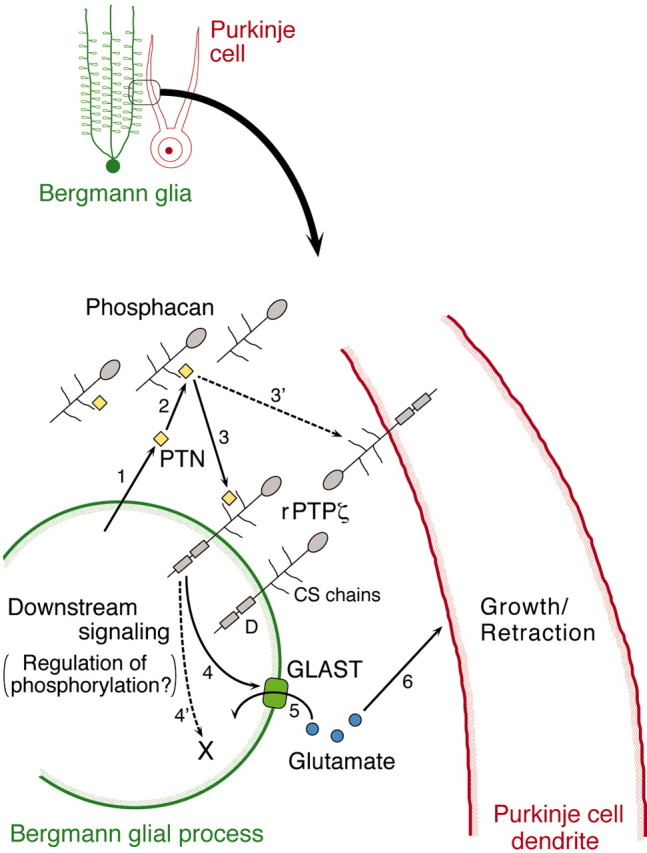
PTPzeta/RPTPbeta, a receptor-type protein tyrosine phosphatase synthesized as a chondroitin sulfate (CS) proteoglycan, uses a heparin-binding growth factor pleiotrophin (PTN) as a ligand, in which the CS portion plays an essential role in ligand binding. Using an organotypic slice culture system, we tested the hypothesis that PTN-PTPzeta signaling is involved in the morphogenesis of Purkinje cell dendrites. An aberrant morphology of Purkinje cell dendrites such as multiple and disoriented primary dendrites was induced in slice cultures by (1) addition of a polyclonal antibody against the extracellular domain of PTPzeta, (2) inhibition of protein tyrosine phosphatase activity, (3) enzymatic removal of the CS chains, (4) addition of exogenous CS chains, and (5) addition of exogenous PTN, all of which disturb PTN-PTPzeta signaling. These treatments also reduced the immunoreactivity to GLAST, a glial glutamate transporter, on Bergmann glial processes. Furthermore, a glutamate transporter inhibitor also induced the abnormal morphogenesis of Purkinje cell dendrites. Altogether, these findings suggest that PTN-PTPzeta signaling regulates the morphogenesis of Purkinje cell dendrites and that the mechanisms underlying that regulation involve the GLAST activity in Bergmann glial processes.
Figures
References
-
- Altman J, Anderson WJ. Experimental reorganization of the cerebellar cortex. I. Morphological effects of eliminating all microneurons with prolonged X-irradiation started at birth. J Comp Neurol. 1972;146:355–406. - PubMed
-
- Amet LEA, Lauri SE, Hienola A, Croll SD, Lu Y, Levorse JM, Prabhakaran B, Taira T, Rauvala H, Vogt TF. Enhanced hippocampal long-term potentiation in mice lacking heparin-binding growth-associated molecule. Mol Cell Neurosci. 2001;17:1014–1024. - PubMed
-
- Armengol J-A, Sotelo C. Early dendritic development of Purkinje cells in the rat cerebellum. A light and electron microscopic study using axonal tracing in “in vitro” slices. Dev Brain Res. 1991;64:95–114. - PubMed
-
- Bandtlow CE, Zimmermann DR. Proteoglycans in the developing brain: new conceptual insights for old proteins. Physiol Rev. 2000;80:1267–1290. - PubMed
-
- Baptista CA, Hatten ME, Blazeski R, Mason CA. Cell-cell interactions influence survival and differentiation of purified Purkinje cells in vitro. Neuron. 1994;12:243–260. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases