

Dopamine modulates cell cycle in the lateral ganglionic eminence
- PMID: 12684471
- PMCID: PMC1201391
- DOI: 10.1523/JNEUROSCI.23-07-02840.2003
Dopamine modulates cell cycle in the lateral ganglionic eminence
Abstract
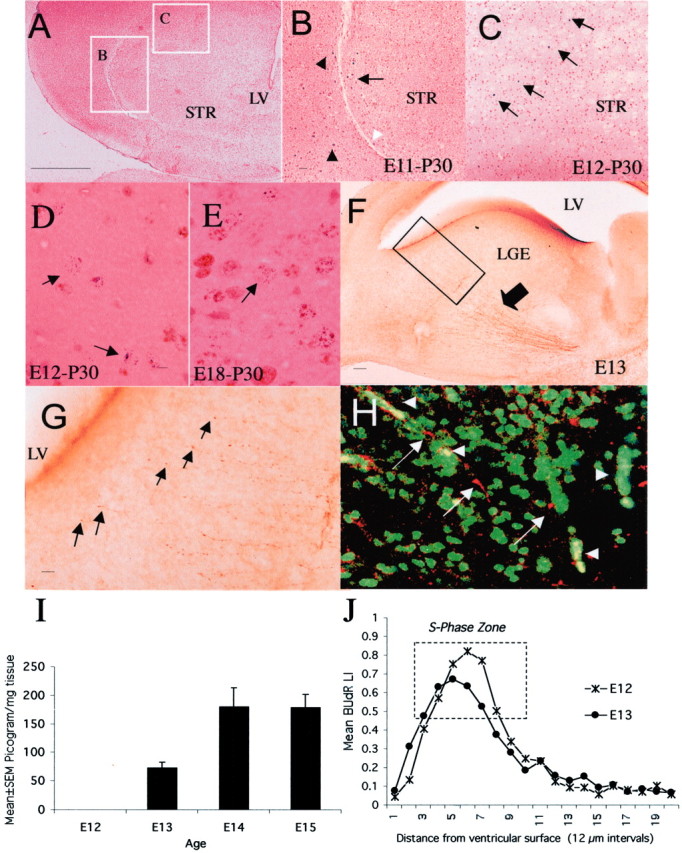
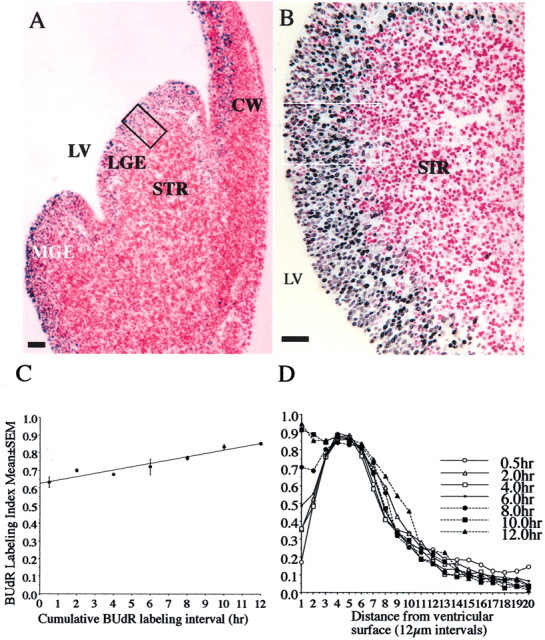
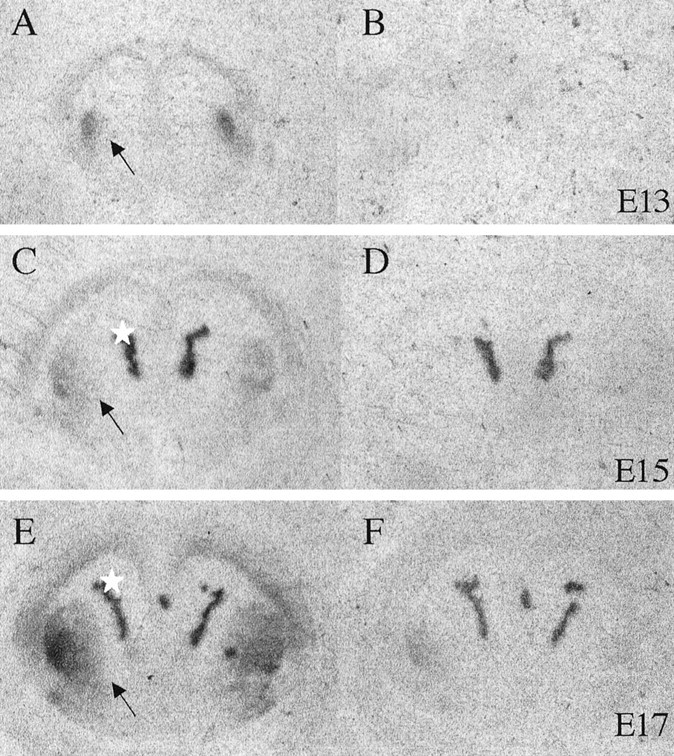
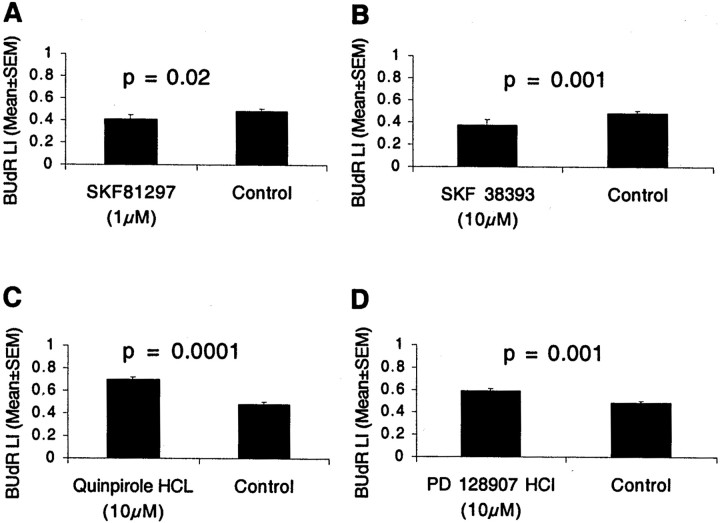
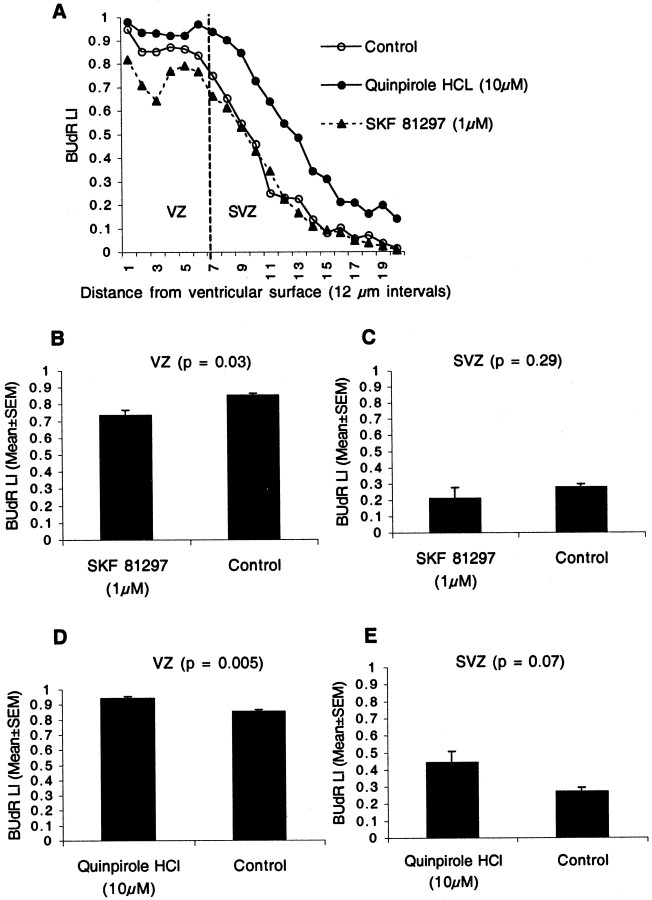
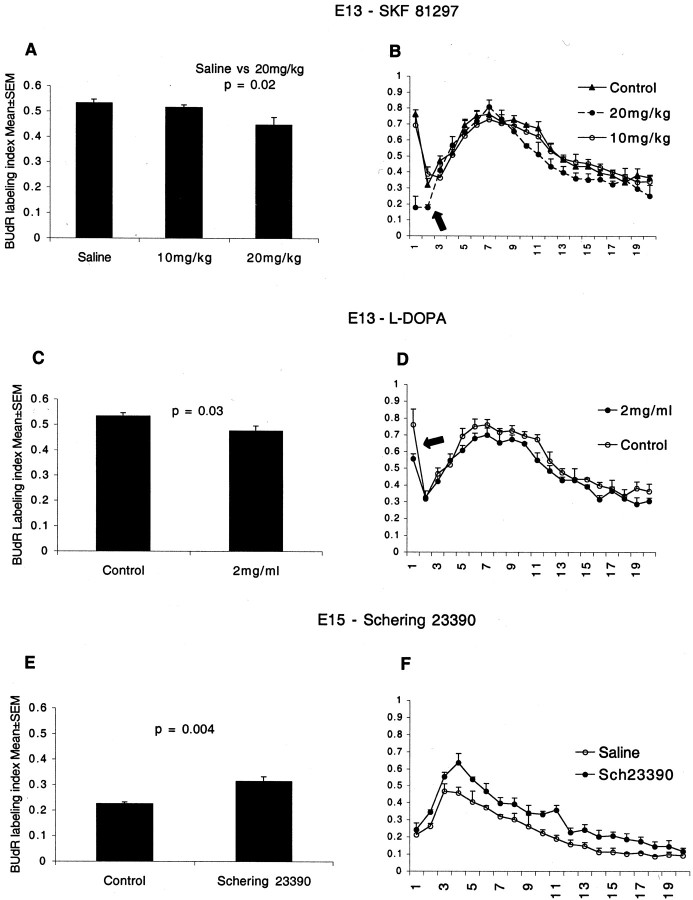
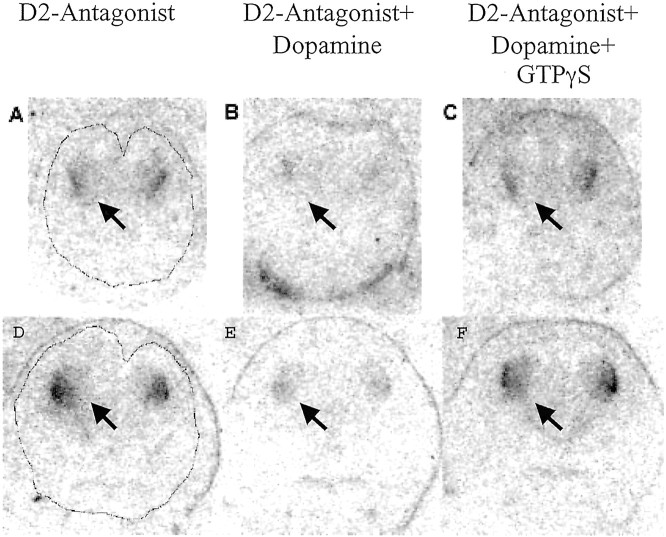
Dopamine is a neuromodulator the functions of which in the regulation of complex behaviors such as mood, motivation, and attention are well known. Dopamine appears in the brain early in the embryonic period when none of those behaviors is robust, raising the possibility that dopamine may influence brain development. The effects of dopamine on specific developmental processes such as neurogenesis are not fully characterized. The neostriatum is a dopamine-rich region of the developing and mature brain. If dopamine influenced neurogenesis, the effects would likely be pronounced in the neostriatum. Therefore, we examined whether dopamine influenced neostriatal neurogenesis by influencing the cell cycle of progenitor cells in the lateral ganglionic eminence (LGE), the neuroepithelial precursor of the neostriatum. We show that dopamine arrives in the LGE via the nigrostriatal pathway early in the embryonic period and that neostriatal neurogenesis progresses in a dopamine-rich milieu. Dopamine D1-like receptor activation reduces entry of progenitor cells from the G(1)- to S-phase of the cell cycle, whereas D2-like receptor activation produces the opposite effects by promoting G(1)- to S-phase entry. D1-like effects are prominent in the ventricular zone, and D2-like effects are prominent in the subventricular zone. The overall effects of dopamine on the cell cycle are D1-like effects, most likely because of the preponderance of D1-like binding sites in the embryonic neostriatum. These data reveal a novel developmental role for dopamine and underscore the relevance of dopaminergic signaling in brain development.
Figures
Similar articles
-
Influence of dopamine on precursor cell proliferation and differentiation in the embryonic mouse telencephalon.Dev Neurosci. 2004 Mar-Aug;26(2-4):229-44. doi: 10.1159/000082140. Dev Neurosci. 2004. PMID: 15711063 Free PMC article.
-
Anatomical and physiological evidence for D1 and D2 dopamine receptor colocalization in neostriatal neurons.Nat Neurosci. 2000 Mar;3(3):226-30. doi: 10.1038/72929. Nat Neurosci. 2000. PMID: 10700253
-
Differential regulation of dopamine D1 and D2 signaling by nicotine in neostriatal neurons.J Neurochem. 2004 Sep;90(5):1094-103. doi: 10.1111/j.1471-4159.2004.02574.x. J Neurochem. 2004. PMID: 15312165
-
Phosphodiesterase 10A inhibitor, MP-10 (PF-2545920), produces greater induction of c-Fos in dopamine D2 neurons than in D1 neurons in the neostriatum.Neuropharmacology. 2015 Dec;99:379-86. doi: 10.1016/j.neuropharm.2015.08.008. Epub 2015 Aug 7. Neuropharmacology. 2015. PMID: 26256420
-
Dopaminergic modulation of neurogenesis in the subventricular zone of the adult brain.Cell Cycle. 2009 Sep 15;8(18):2888-94. doi: 10.4161/cc.8.18.9512. Epub 2009 Sep 14. Cell Cycle. 2009. PMID: 19713754 Review.
Cited by
-
Thalamocortical functional connectivity and behavioral disruptions in neonates with prenatal cocaine exposure.Neurotoxicol Teratol. 2016 Jul-Aug;56:16-25. doi: 10.1016/j.ntt.2016.05.009. Epub 2016 May 27. Neurotoxicol Teratol. 2016. PMID: 27242332 Free PMC article.
-
Neurotransmitter-mediated control of neurogenesis in the adult vertebrate brain.Development. 2013 Jun;140(12):2548-61. doi: 10.1242/dev.088005. Development. 2013. PMID: 23715548 Free PMC article. Review.
-
A transgenic mouse model of neuroepithelial cell specific inducible overexpression of dopamine D1-receptor.Neuroscience. 2010 Oct 27;170(3):961-70. doi: 10.1016/j.neuroscience.2010.07.036. Epub 2010 Jul 29. Neuroscience. 2010. PMID: 20674683 Free PMC article.
-
Cocaine exposure decreases GABA neuron migration from the ganglionic eminence to the cerebral cortex in embryonic mice.Cereb Cortex. 2004 Jun;14(6):665-75. doi: 10.1093/cercor/bhh027. Epub 2004 Mar 28. Cereb Cortex. 2004. PMID: 15054047 Free PMC article.
-
Reduced Prefrontal Gyrification in Carriers of the Dopamine D4 Receptor 7-Repeat Allele With Attention Deficit/Hyperactivity Disorder: A Preliminary Report.Front Psychiatry. 2019 Apr 25;10:235. doi: 10.3389/fpsyt.2019.00235. eCollection 2019. Front Psychiatry. 2019. PMID: 31105599 Free PMC article.
References
-
- Akbarian S, Kim JJ, Potkin SG, Hagman JO, Tafazzoli A, Bunney WE, Jr, Jones EG. Gene expression for glutamic acid decarboxylase is reduced without loss of neurons in prefrontal cortex of schizophrenics. Arch Gen Psychiatry. 1995;52:258–266. - PubMed
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JLR. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Benes FM. Emerging principles of altered neural circuitry in schizophrenia. Brain Res Brain Res Rev. 2000;31:251–269. - PubMed
-
- Benes FM, Vincent SL, Marie A, Kahn Y. Up-regulation of GABA receptor binding on neurons of the prefrontal cortex in schizophrenic subjects. Neuroscience. 1996;75:1021–1031. - PubMed
-
- Bhide PG. Cell cycle kinetics in the embryonic mouse corpus striatum. J Comp Neurol. 1996;374:506–522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases