

Xylulose 5-phosphate mediates glucose-induced lipogenesis by xylulose 5-phosphate-activated protein phosphatase in rat liver
- PMID: 12684532
- PMCID: PMC154306
- DOI: 10.1073/pnas.0730817100
Xylulose 5-phosphate mediates glucose-induced lipogenesis by xylulose 5-phosphate-activated protein phosphatase in rat liver
Abstract
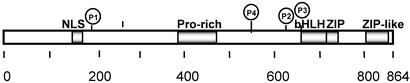
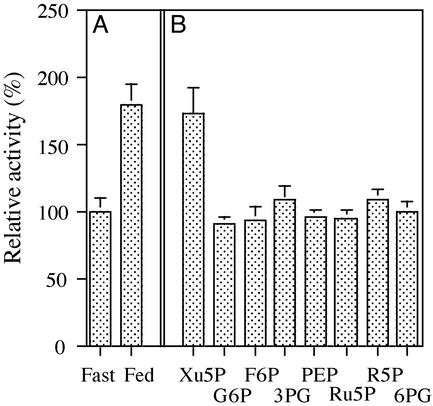
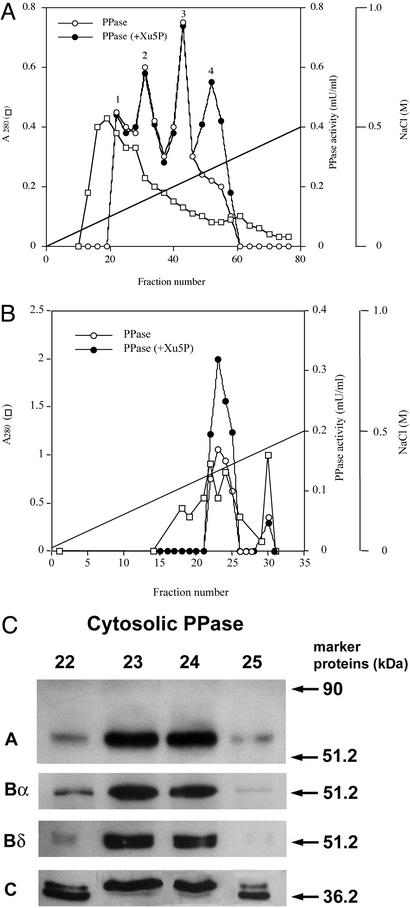
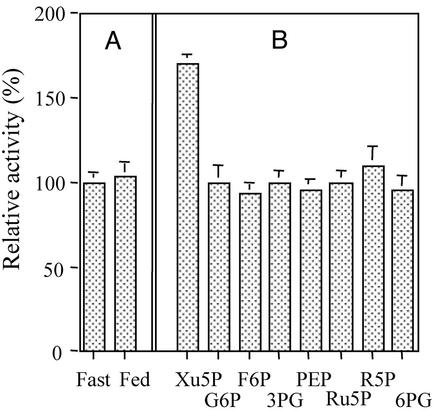
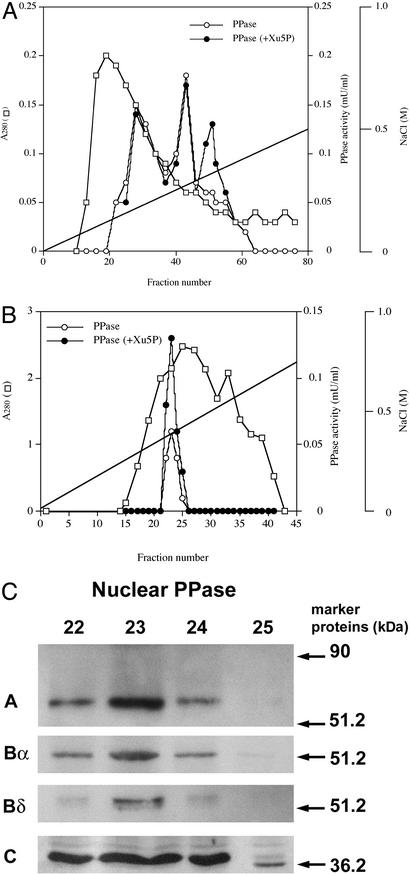
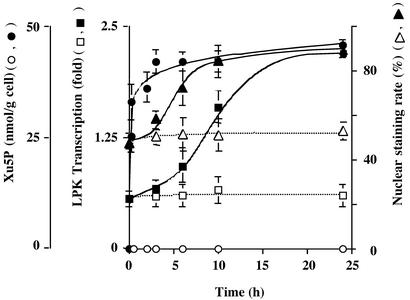
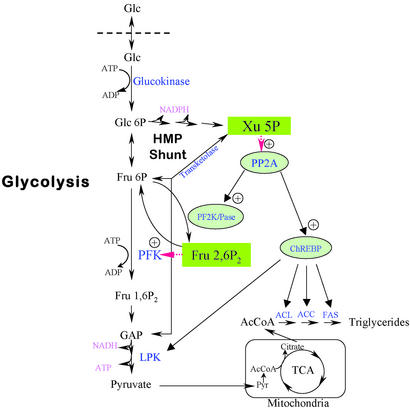
Carbohydrate-responsive element binding protein (ChREBP) is a transcription factor that activates lipogenic genes in liver in response to excess carbohydrate in the diet. ChREBP is regulated in a reciprocal manner by glucose and cAMP. cAMP-dependent protein kinase (protein kinase A) phosphorylates two physiologically important sites in ChREBP, Ser-196, which is located near nuclear localization signal sequence (NLS), and Thr-666, within the basic helix-loop-helix (bHLH) site, resulting in inactivation of nuclear translocation of ChREBP and of the DNA-binding activity, respectively. We demonstrate here that crude cytosolic extracts from livers of rats fed a high carbohydrate diet contained protein phosphatase (PPase) activity that dephosphorylated a peptide containing Ser-196, whereas a PPase in the nuclear extract catalyzed dephosphorylation of Ser-568 and Thr-666. All these PPases are activated specifically by xylulose 5-phosphate (Xu5P), but not by other sugar phosphates. Furthermore, addition of Xu5P elevated PPase activity to the level observed in extracts of fed liver cells. These partially purified PPases were characterized as PP2A-AB delta C by immunoblotting with specific antibodies. These results suggest that (ia) Xu5P-dependent PPase is responsible for activation of transcription of the L-type pyruvate kinase gene and lipogenic enzyme genes, and (ii) Xu5P is the glucose signaling compound. Thus, we propose that the same Xu5P-activated PPase controls both acute and long-term regulation of glucose metabolism and fat synthesis.
Figures
Comment in
-
A humble hexose monophosphate pathway metabolite regulates short- and long-term control of lipogenesis.Proc Natl Acad Sci U S A. 2003 May 13;100(10):5578-80. doi: 10.1073/pnas.1132039100. Epub 2003 Apr 29. Proc Natl Acad Sci U S A. 2003. PMID: 12721358 Free PMC article. Review. No abstract available.
References
-
- Girard J, Ferre P, Foufelle F. Annu Rev Nutr. 1997;17:325–352. - PubMed
-
- Towle H C, Kaytor E N, Shih H M. Annu Rev Nutr. 1997;17:405–433. - PubMed
-
- Nishimura M, Fedorov S, Uyeda K. J Biol Chem. 1994;269:26100–26106. - PubMed
-
- Towle H C. J Biol Chem. 1995;270:23235–23238. - PubMed
-
- Vaulont S, Vasseur-Cognet M, Kahn A. J Biol Chem. 2000;275:31555–31558. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
