

Expression and regulation of the Na+-K+-2Cl- cotransporter NKCC1 in the normal and CFTR-deficient murine colon
- PMID: 12692180
- PMCID: PMC2342946
- DOI: 10.1113/jphysiol.2002.030205
Expression and regulation of the Na+-K+-2Cl- cotransporter NKCC1 in the normal and CFTR-deficient murine colon
Abstract
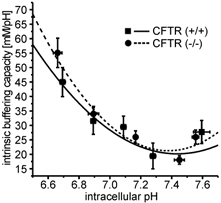
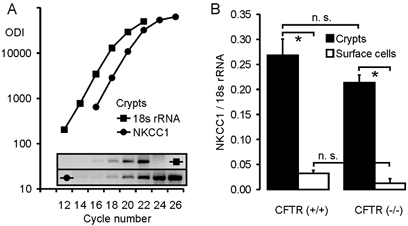
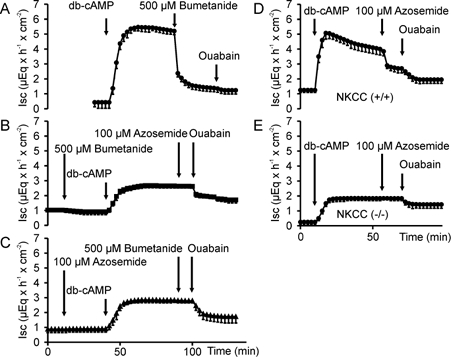
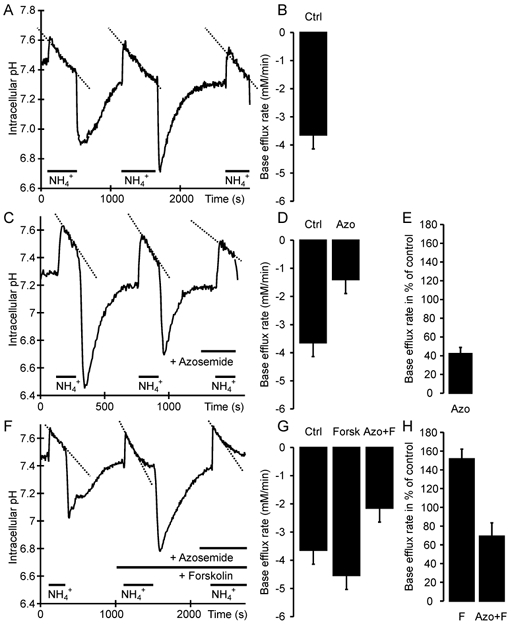
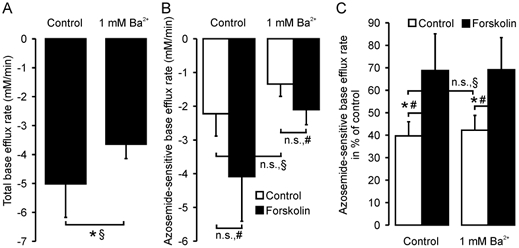
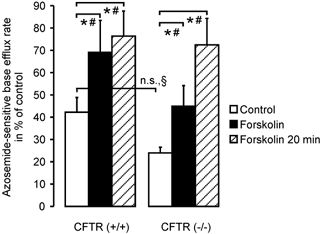
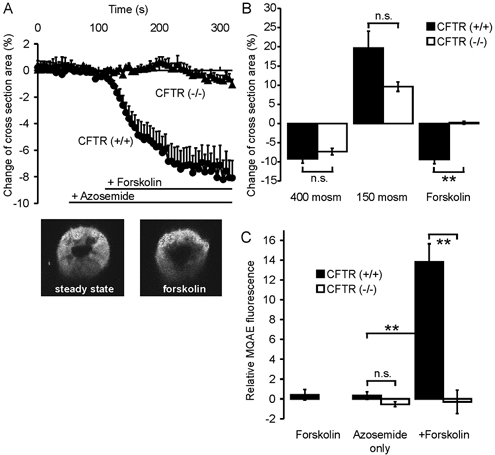
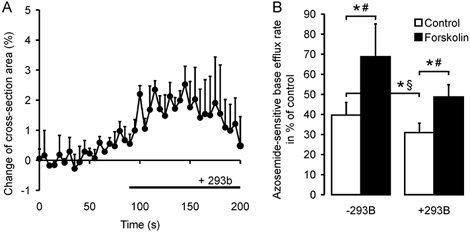
Defective regulation and/or reduced expression of the Na+-K+-2Cl- cotransporter NKCC1 may contribute to the severe secretory defect that is observed in cystic fibrosis, but data concerning the expression and function of NKCC1 in cystic fibrosis transmembrane conductance regulator (CFTR)-deficient cells are equivocal. We therefore investigated NKCC1 mRNA expression, Na+-K+-2Cl- cotransport activity and regulation by cAMP in crypts isolated from the proximal colon of CFTR-containing (CFTR (+/+)) and CFTR-deficient (CFTR (-/-)) mice. mRNA expression levels were determined by semiquantitative PCR, transport rates were measured fluorometrically in 2',7'-bis(carboxyethyl)-5(6)-carboxyfluorescein acetomethylester (BCECF)-loaded crypts, cytoplasmic volume changes were assessed by confocal microscopy, and [Cl-]i changes were examined by N-(ethoxycarbonylmethyl)-6-methoxyquinolinium bromide (MQAE) quenching. NKCC1 mRNA expression levels were not significantly reduced in CFTR (-/-) crypts compared to controls. Azosemide-sensitive NH4+ influx (used as a measure of Na+-K+-2Cl- cotransport) was 2.23 +/- 0.72 vs. 1.56 +/- 0.16 mM min-1, and increased by 63.6 % in (+/+) and 87.3 % in (-/-) crypts upon stimulation for 5 min with forskolin. After 20 min of stimulation with forskolin, the Na+-K+-2Cl- cotransport rates in (-/-) and (+/+) crypts were identical. Crypt cross-sectional area and [Cl-]i decreased only in (+/+) crypts upon stimulation. In conclusion, normal NKCC1 expression levels, somewhat reduced Na+-K+-2Cl- cotransport rates, but preserved activation by cAMP were found in colonic crypts from CFTR (-/-) mice, ruling out a severe dysfunction of the Na+-K+-2Cl- cotransporter in the CF intestine. Furthermore, these studies establish the existence of a direct, cell-volume- and [Cl-]i-independent activation of colonic NKCC1 by an increase in intracellular cAMP.
Figures
References
-
- Bachmann O, Rossmann H, Berger UV, Colledge WH, Ratcliff R, Evans MJ, Gregor M, Seidler U. cAMP-mediated regulation of murine intestinal/pancreatic Na+/HCO3− cotransporter subtype pNBC1. Am J Physiol. 2003;284:G37–45. - PubMed
-
- Bachmann O, Sonnentag T, Siegel WK, Lamprecht G, Weichert A, Gregor M, Seidler U. Different acid secretagogues activate different Na+/H+ exchanger isoforms in rabbit parietal cells. Am J Physiol. 1998;275:G1085–1093. - PubMed
-
- Barrett KE, Keely SJ. Chloride secretion by the intestinal epithelium: molecular basis and regulatory aspects. Annu Rev Physiol. 2000;62:535–572. - PubMed
-
- Boyarsky G, Ganz MB, Sterzel RB, Boron WF. pH regulation in single glomerular mesangial cells. I. Acid extrusion in absence and presence of HCO3−. Am J Physiol. 1988;255:C844–856. - PubMed
-
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous