

Conformational changes, plasma membrane penetration, and infection by human rhinovirus type 2: role of receptors and low pH
- PMID: 12692239
- PMCID: PMC153956
- DOI: 10.1128/jvi.77.9.5370-5377.2003
Conformational changes, plasma membrane penetration, and infection by human rhinovirus type 2: role of receptors and low pH
Abstract
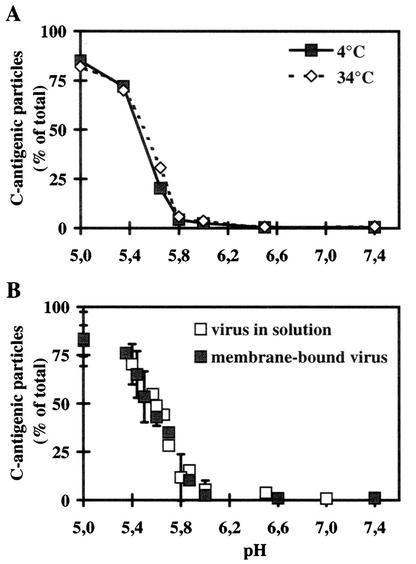
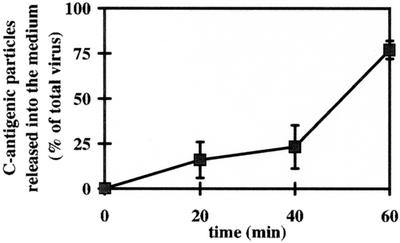
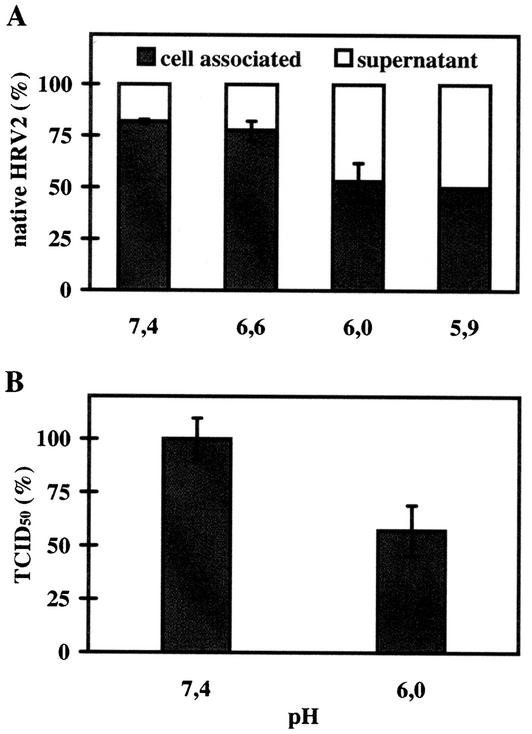
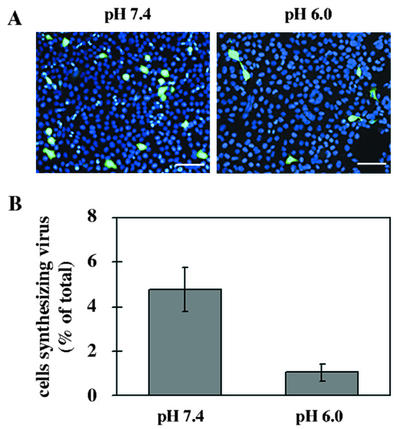
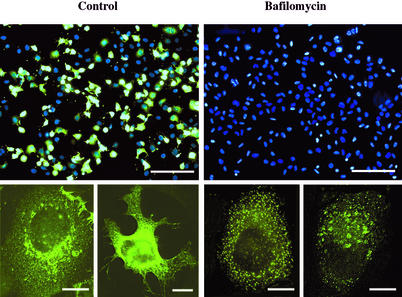
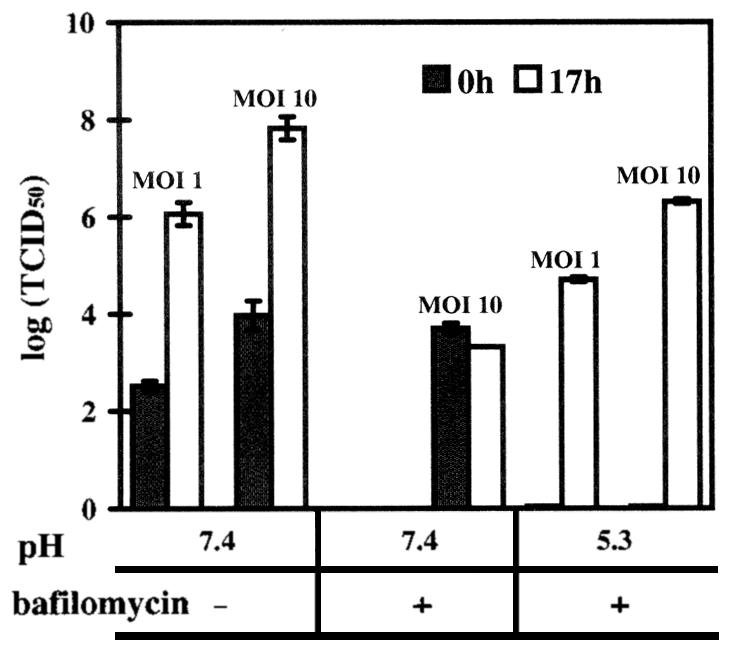
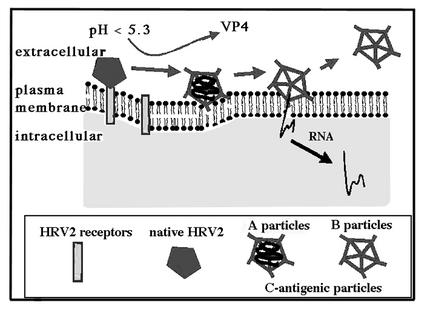
Human rhinovirus type 2 (HRV2) is internalized by members of the low-density lipoprotein (LDL) receptor (LDLR) family. It then progresses into late endosomes, where it undergoes conversion from D- to C-antigenicity at pH < 5.6. Upon uncoating, the viral RNA is transferred into the cytoplasm across the endsosomal membrane. However, C-antigenic particles fail to attach to LDLR; this raised the question of whether the virus remains attached to the receptors and is carried to late compartments or rather falls off at the higher pH in early endosomes. We therefore determined the pH dependence of virus-receptor dissociation and virus conversion to C-antigen under conditions preventing endocytosis. (35)S-HRV2 was attached to HeLa cells at 4 degrees C and incubated in buffers of pH 7.4 to 5.0; levels of native virus and C-antigenic particles remaining cell associated or having been released into the medium were determined by immunoprecipitation. At pH 6.0, HRV2 was readily released from plasma membrane receptors in its native form, whereas at pH < or = 5.4, it was entirely converted to C-antigen, which, however, only dissociated from the surface upon prolonged incubation. The antigenic conversion occurred at the same pH regardless of whether HRV2 was free in solution or bound to its receptors. These data suggest that, in vivo, the virus is no longer bound to its receptors when the antigenic conversion and uncoating occur in more acidic late endosomes. When virus was bound to HeLa cells at 4 degrees C, converted into C-antigen by exposure to pH 5.3, and subsequently warmed to 34 degrees C in the presence of bafilomycin (to prevent endosomal uncoating), viral de novo synthesis was detected. This study demonstrates for the first time that a nonenveloped virus such as HRV2 can infect from the plasma membrane when artificially exposed to low pH. This implies that the viral RNA can gain access to the cytoplasm from the plasma membrane.
Figures
References
-
- Bayer, N., E. Prchla, M. Schwab, D. Blaas, and R. Fuchs. 1999. Human rhinovirus HRV14 uncoats from early endosomes in the presence of bafilomycin. FEBS Lett. 463:175-178. - PubMed
-
- Blake, K., and S. O'Connell. 1993. Virus culture, p. 81-122. In D. R. Harper (ed.), Virology Labfax. Blackwell Scientific Publications, London, United Kingdom.
-
- Brown, M. S., J. Herz, and J. L. Goldstein. 1997. LDL receptor structure: calcium cages, acid baths and recycling receptors. Nature 388:629-630. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
