

NF2 deficiency promotes tumorigenesis and metastasis by destabilizing adherens junctions
- PMID: 12695331
- PMCID: PMC196046
- DOI: 10.1101/gad.1054603
NF2 deficiency promotes tumorigenesis and metastasis by destabilizing adherens junctions
Abstract
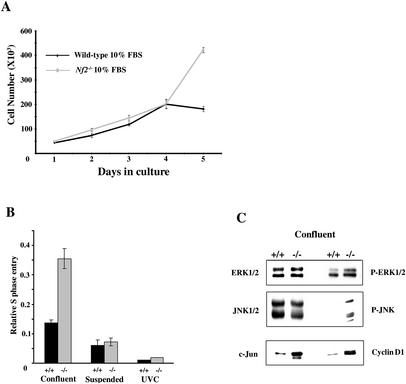
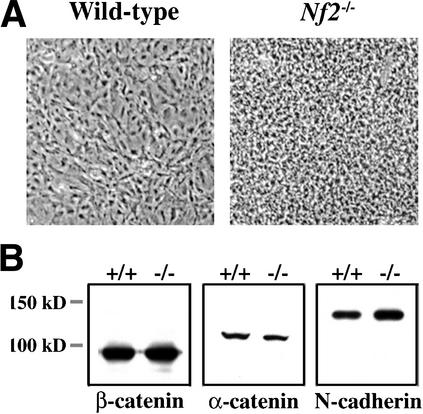
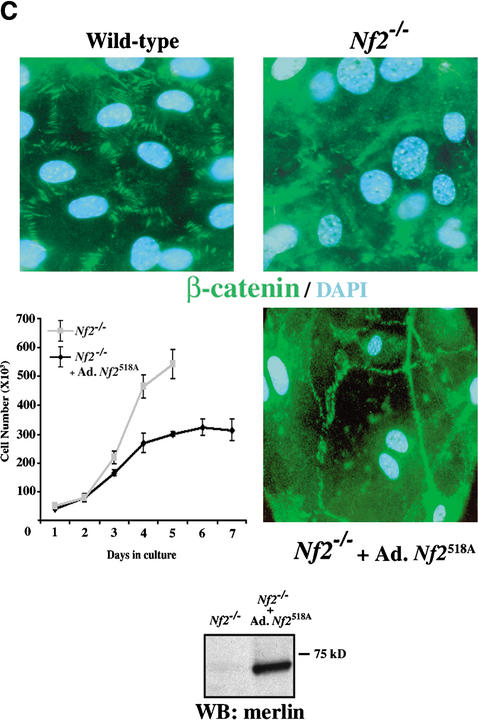
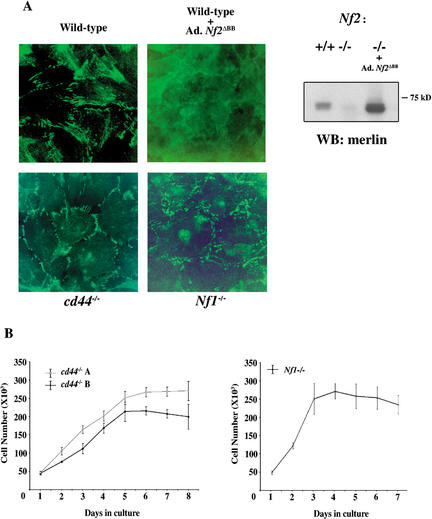
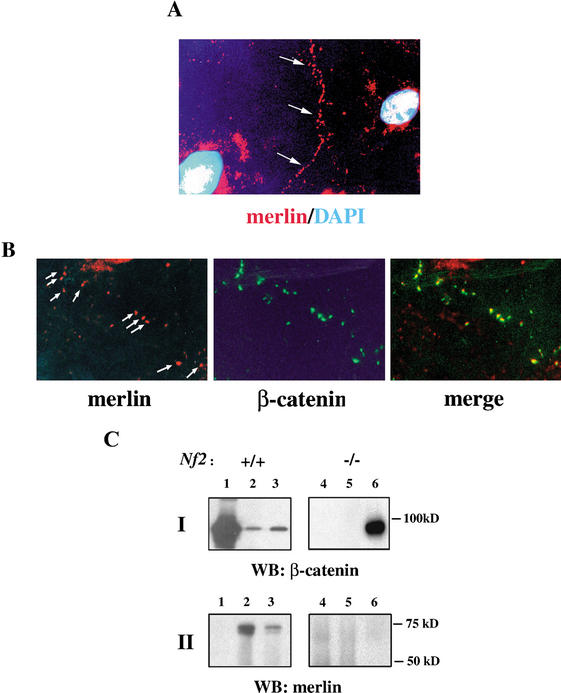
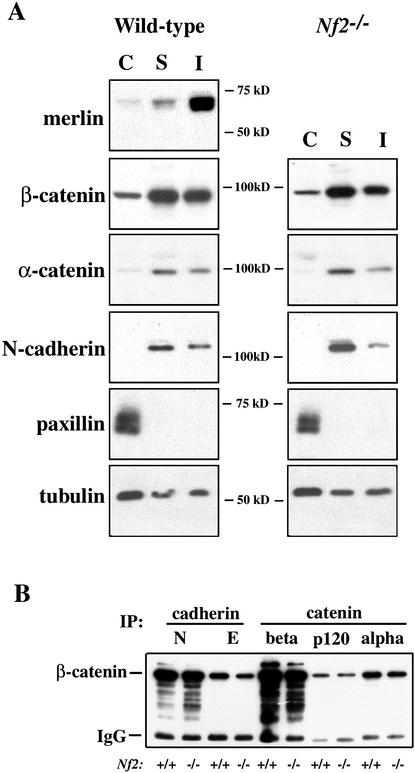
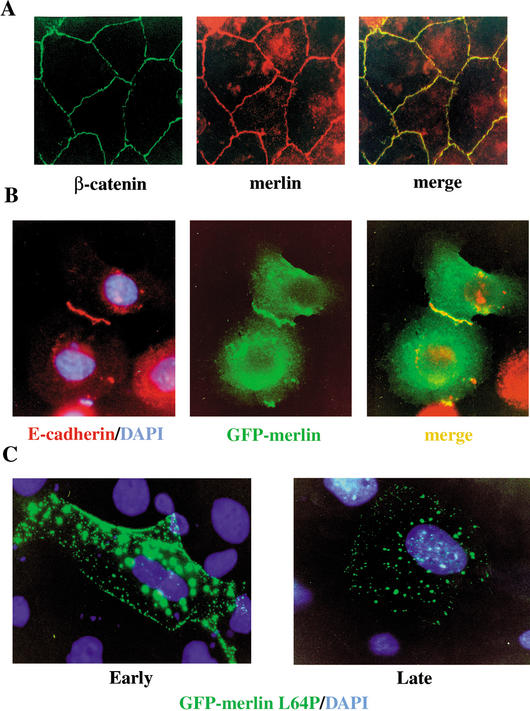
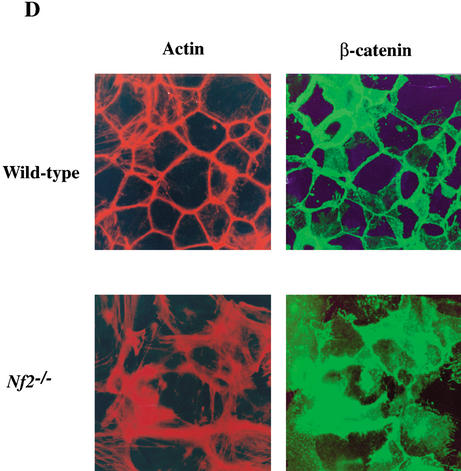
Mutation of the Neurofibromatosis 2 (NF2) tumor suppressor gene leads to cancer development in humans and mice. Recent studies suggest that Nf2 loss also contributes to tumor metastasis. The Nf2-encoded protein, merlin, is related to the ERM (ezrin, radixin, and moesin) family of membrane:cytoskeleton-associated proteins. However, the cellular mechanism whereby merlin controls cell proliferation from this location is not known. Here we show that the major cellular consequence of Nf2 deficiency in primary cells is an inability to undergo contact-dependent growth arrest and to form stable cadherin-containing cell:cell junctions. Merlin colocalizes and interacts with adherens junction (AJ) components in confluent wild-type cells, suggesting that the lack of AJs and contact-dependent growth arrest in Nf2(-/-) cells directly results from the absence of merlin at sites of cell:cell contact. Our studies indicate that merlin functions as a tumor and metastasis suppressor by controlling cadherin-mediated cell:cell contact.
Figures
References
-
- Bernards R, Weinberg RA. A progression puzzle. Nature. 2002;418:823. - PubMed
-
- Bretscher A, Edwards K, Fehon RG. ERM proteins and merlin: Integrators at the cell cortex. Nat Rev Mol Cell Biol. 2002;3:586–599. - PubMed
-
- Cavallaro U, Christofori G. Cell adhesion in tumor invasion andmetastasis: Loss of the glue is not enough. Biochim Biophys Acta. 2001;1552:39–45. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous