

The Hsp70 and TRiC/CCT chaperone systems cooperate in vivo to assemble the von Hippel-Lindau tumor suppressor complex
- PMID: 12697815
- PMCID: PMC153194
- DOI: 10.1128/MCB.23.9.3141-3151.2003
The Hsp70 and TRiC/CCT chaperone systems cooperate in vivo to assemble the von Hippel-Lindau tumor suppressor complex
Abstract
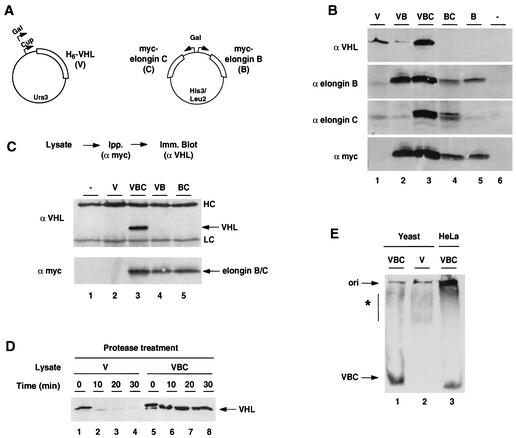
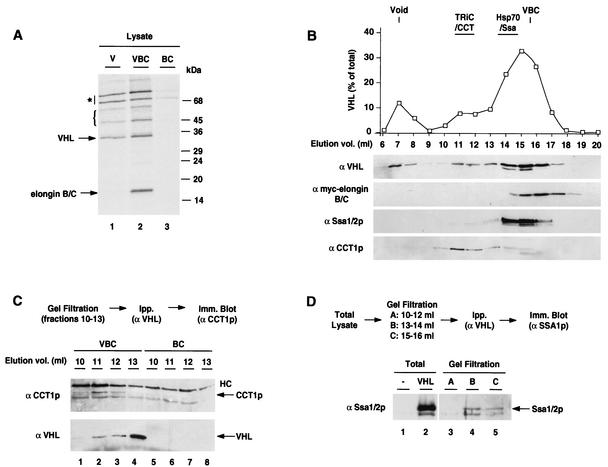
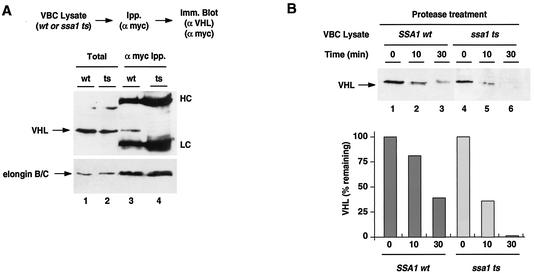
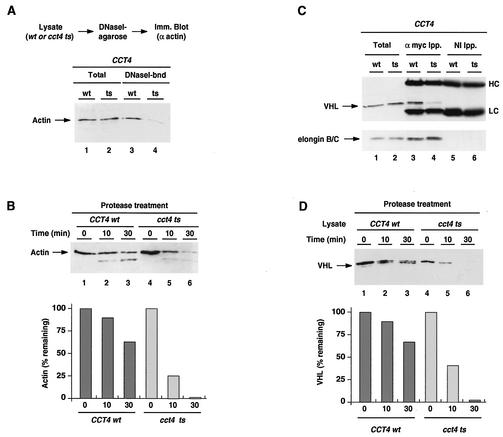
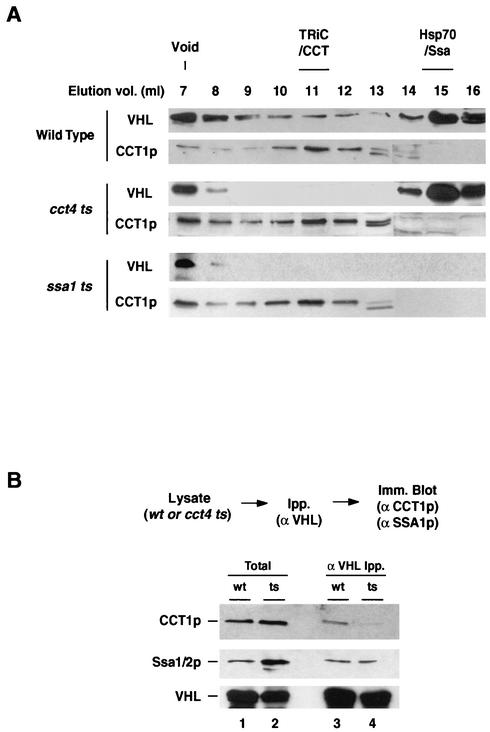
The degree of cooperation and redundancy between different chaperones is an important problem in understanding how proteins fold in the cell. Here we use the yeast Saccharomyces cerevisiae as a model system to examine in vivo the chaperone requirements for assembly of the von Hippel-Lindau protein (VHL)-elongin BC (VBC) tumor suppressor complex. VHL and elongin BC expressed in yeast assembled into a correctly folded VBC complex that resembles the complex from mammalian cells. Unassembled VHL did not fold and remained associated with the cytosolic chaperones Hsp70 and TRiC/CCT, in agreement with results from mammalian cells. Analysis of the folding reaction in yeast strains carrying conditional chaperone mutants indicates that incorporation of VHL into VBC requires both functional TRiC and Hsp70. VBC assembly was defective in cells carrying either a temperature-sensitive ssa1 gene as their sole source of cytosolic Hsp70/SSA function or a temperature-sensitive mutation in CCT4, a subunit of the TRiC/CCT complex. Analysis of the VHL-chaperone interactions in these strains revealed that the cct4ts mutation decreased binding to TRiC but did not affect the interaction with Hsp70. In contrast, loss of Hsp70 function disrupted the interaction of VHL with both Hsp70 and TRiC. We conclude that, in vivo, folding of some polypeptides requires the cooperation of Hsp70 and TRiC and that Hsp70 acts to promote substrate binding to TRiC.
Figures
Similar articles
-
Formation of the VHL-elongin BC tumor suppressor complex is mediated by the chaperonin TRiC.Mol Cell. 1999 Dec;4(6):1051-61. doi: 10.1016/s1097-2765(00)80233-6. Mol Cell. 1999. PMID: 10635329
-
Diverse effects of mutations in exon II of the von Hippel-Lindau (VHL) tumor suppressor gene on the interaction of pVHL with the cytosolic chaperonin and pVHL-dependent ubiquitin ligase activity.Mol Cell Biol. 2002 Mar;22(6):1947-60. doi: 10.1128/MCB.22.6.1947-1960.2002. Mol Cell Biol. 2002. PMID: 11865071 Free PMC article.
-
Tumorigenic mutations in VHL disrupt folding in vivo by interfering with chaperonin binding.Mol Cell. 2003 Nov;12(5):1213-24. doi: 10.1016/s1097-2765(03)00423-4. Mol Cell. 2003. PMID: 14636579
-
The von Hippel-Lindau tumor suppressor protein.Curr Opin Genet Dev. 2001 Feb;11(1):27-34. doi: 10.1016/s0959-437x(00)00152-0. Curr Opin Genet Dev. 2001. PMID: 11163147 Review.
-
The role of molecular chaperones in human misfolding diseases.FEBS Lett. 2009 Aug 20;583(16):2647-53. doi: 10.1016/j.febslet.2009.04.029. Epub 2009 Apr 23. FEBS Lett. 2009. PMID: 19393652 Review.
Cited by
-
Regulation of GPCR expression through an interaction with CCT7, a subunit of the CCT/TRiC complex.Mol Biol Cell. 2016 Dec 1;27(24):3800-3812. doi: 10.1091/mbc.E16-04-0224. Epub 2016 Oct 5. Mol Biol Cell. 2016. PMID: 27708139 Free PMC article.
-
Molecular chaperones: Guardians of tumor suppressor stability and function.Oncotarget. 2024 Oct 1;15:679-696. doi: 10.18632/oncotarget.28653. Oncotarget. 2024. PMID: 39352796 Free PMC article. Review.
-
Sgt1 associates with Hsp90: an initial step of assembly of the core kinetochore complex.Mol Cell Biol. 2004 Sep;24(18):8069-79. doi: 10.1128/MCB.24.18.8069-8079.2004. Mol Cell Biol. 2004. PMID: 15340069 Free PMC article.
-
CCT6A knockdown suppresses osteosarcoma cell growth and Akt pathway activation in vitro.PLoS One. 2022 Dec 30;17(12):e0279851. doi: 10.1371/journal.pone.0279851. eCollection 2022. PLoS One. 2022. PMID: 36584147 Free PMC article.
-
Glycosylation of Skp1 promotes formation of Skp1-cullin-1-F-box protein complexes in dictyostelium.Mol Cell Proteomics. 2015 Jan;14(1):66-80. doi: 10.1074/mcp.M114.044560. Epub 2014 Oct 23. Mol Cell Proteomics. 2015. PMID: 25341530 Free PMC article.
References
-
- Aso, T., and M. N. Conrad. 1997. Molecular cloning of DNAs encoding the regulatory subunits of elongin from Saccharomyces cerevisiae and Drosophila melanogaster. Biochem. Biophys. Res. Commun. 241:334-340. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1992. Current protocols in molecular biology, vol. 1. Wiley-Interscience, New York, N.Y.
-
- Bazin, R., and R. Lemieux. 1989. Increased proportion of B-cell hybridomas secreting monoclonal antibodies of desired specificity in cultures containing macrophage-derived hybridoma growth factor (IL-6). J. Immunol. Methods 116:245-249. - PubMed
-
- Boorstein, W. R., T. Ziegelhoffer, and E. A. Craig. 1994. Molecular evolution of the Hsp70 multigene family. J. Mol. Evol. 38:1-17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous