

Substrate specificity of the nonribosomal peptide synthetase PvdD from Pseudomonas aeruginosa
- PMID: 12700264
- PMCID: PMC154398
- DOI: 10.1128/JB.185.9.2848-2855.2003
Substrate specificity of the nonribosomal peptide synthetase PvdD from Pseudomonas aeruginosa
Abstract
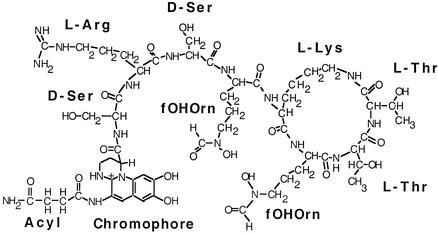
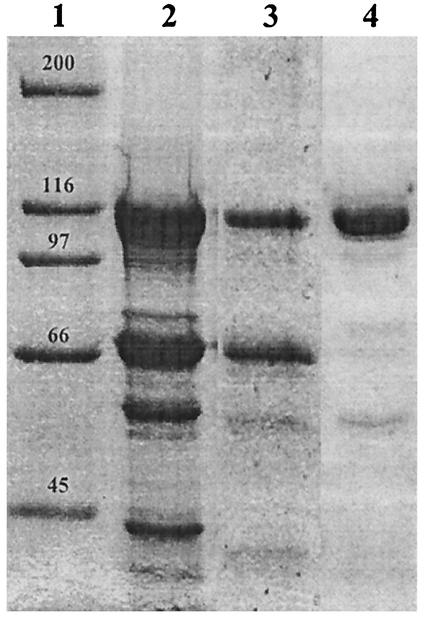
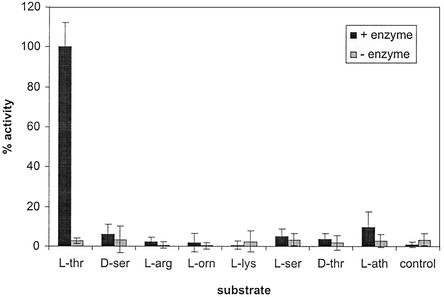
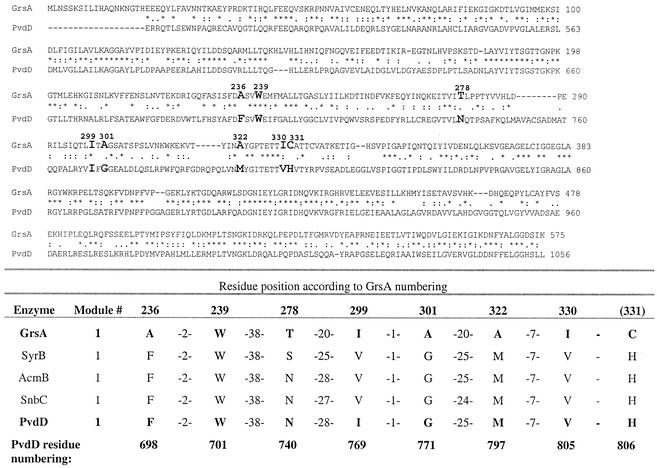
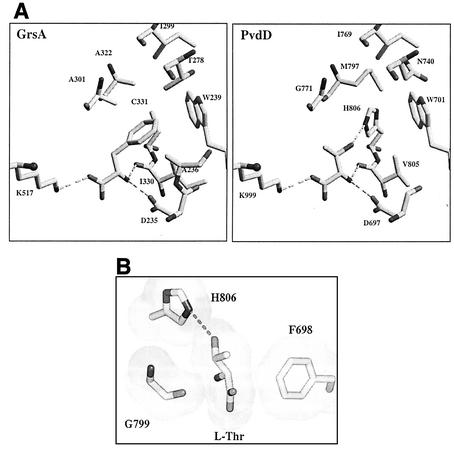
Pseudomonas aeruginosa PAO1 secretes a siderophore, pyoverdine(PAO), which contains a short peptide attached to a dihydroxyquinoline moiety. Synthesis of this peptide is thought to be catalyzed by nonribosomal peptide synthetases, one of which is encoded by the pvdD gene. The first module of pvdD was overexpressed in Escherichia coli, and the protein product was purified. L-Threonine, one of the amino acid residues in pyoverdine(PAO), was an effective substrate for the recombinant protein in ATP-PP(i) exchange assays, showing that PvdD has peptide synthetase activity. Other amino acids, including D-threonine, L-serine, and L-allo-threonine, were not effective substrates, indicating that PvdD has a high degree of substrate specificity. A three-dimensional modeling approach enabled us to identify amino acids that are likely to be critical in determining the substrate specificity of PvdD and to explore the likely basis of the high substrate selectivity. The approach described here may be useful for analysis of other peptide synthetases.
Figures
References
-
- Abdallah, M. A. 1991. Pyoverdines and pseudobactins, p. 139-153. In G. Winkelmann (ed.), CRC handbook of microbial iron chelates. CRC Press, Boca Raton, Fla.
-
- Blackwell, J. R., and R. Horgan. 1991. A novel strategy for production of a highly expressed recombinant protein in an active form. FEBS Lett. 295:10-12. - PubMed
-
- Buchberger, A., and B. Bukau. 1997. Escherichia coli DnaK, p. 22-24. In M. J. Gething (ed.), Guidebook to molecular chaperones and protein-folding catalysts. Sambrook and Tooze, Oxford University Press, Oxford, United Kingdom.
-
- Challis, G. L., J. Ravel, and C. A. Townsend. 2000. Predictive, structure-based model of amino acid recognition by nonribosomal peptide synthetase adenylation domains. Chem. Biol. 7:211-224. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
