

The YSIRK-G/S motif of staphylococcal protein A and its role in efficiency of signal peptide processing
- PMID: 12700270
- PMCID: PMC154403
- DOI: 10.1128/JB.185.9.2910-2919.2003
The YSIRK-G/S motif of staphylococcal protein A and its role in efficiency of signal peptide processing
Abstract
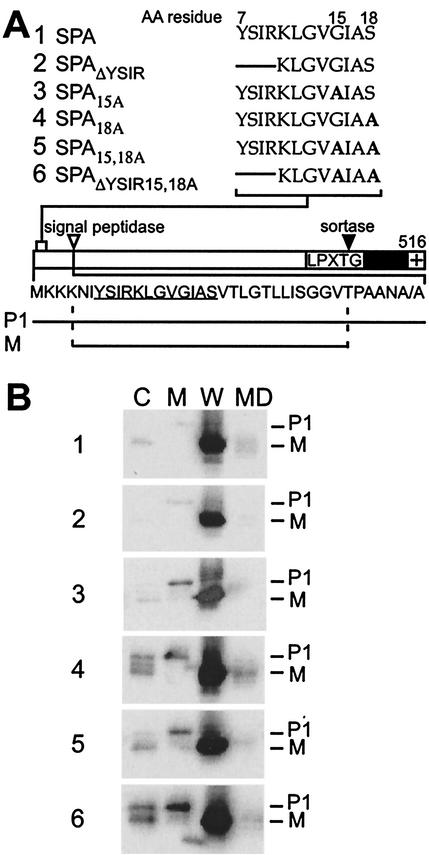
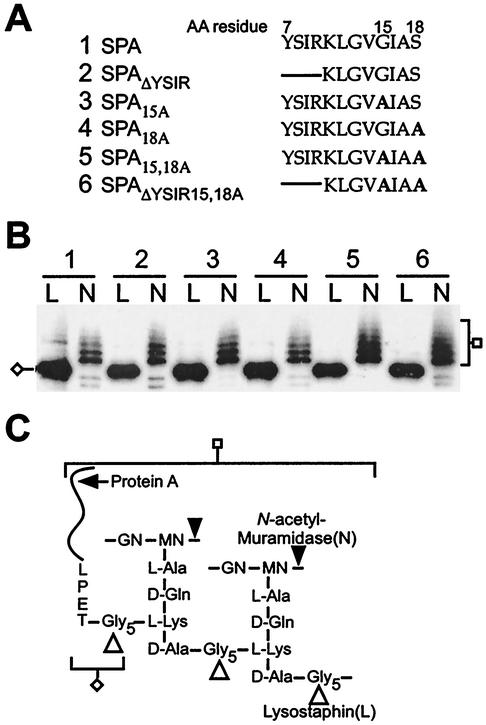
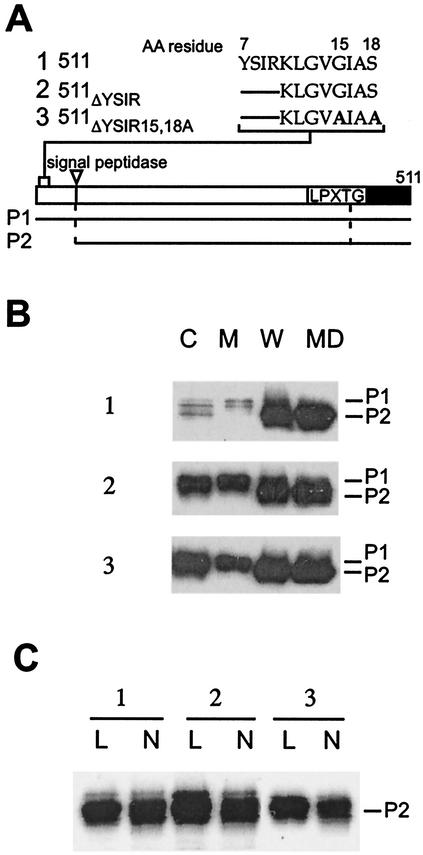
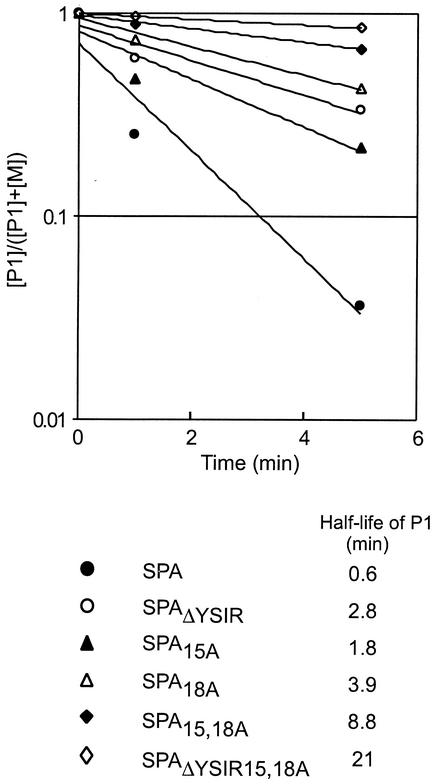
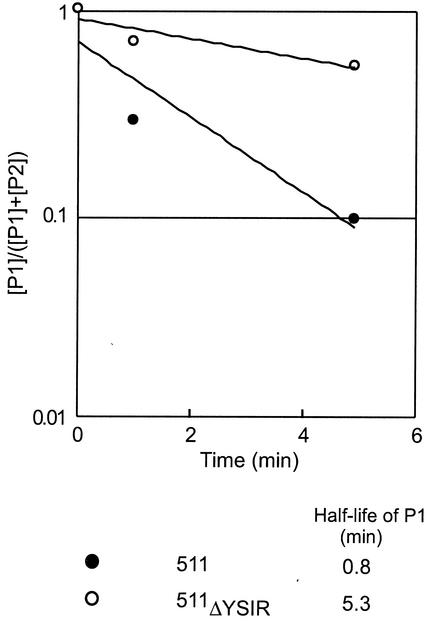
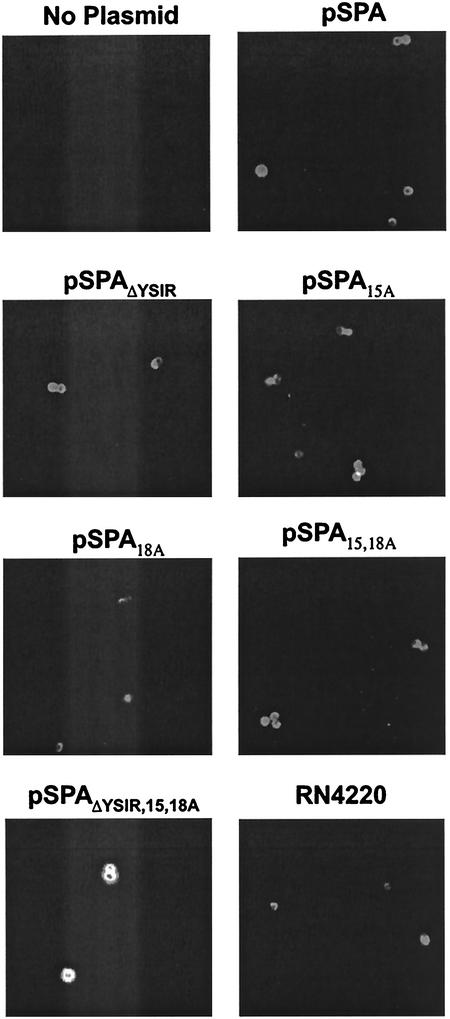
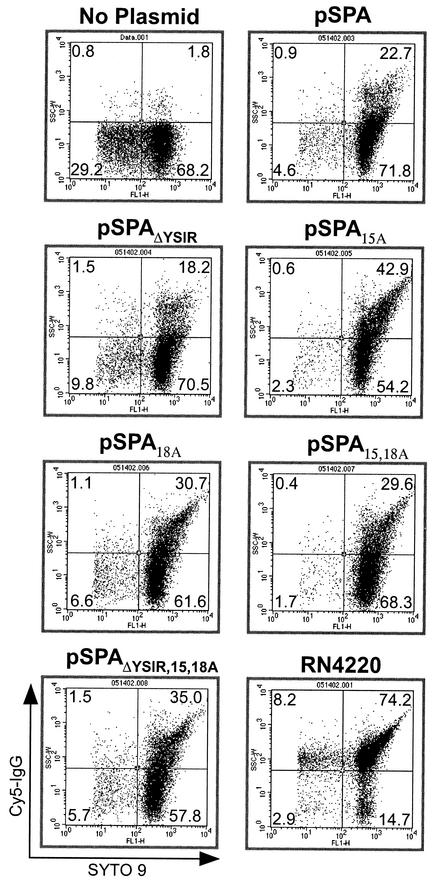
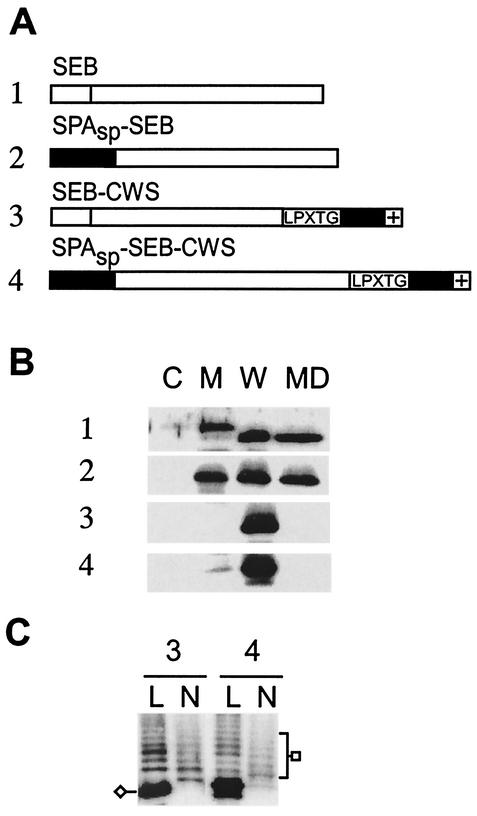
Many surface proteins of pathogenic gram-positive bacteria are linked to the cell wall envelope by a mechanism requiring a C-terminal sorting signal with an LPXTG motif. Surface proteins of Streptococcus pneumoniae harbor another motif, YSIRK-G/S, which is positioned within signal peptides. The signal peptides of some, but not all, of the 20 surface proteins of Staphylococcus aureus carry a YSIRK-G/S motif, whereas those of surface proteins of Listeria monocytogenes and Bacillus anthracis do not. To determine whether the YSIRK-G/S motif is required for the secretion or cell wall anchoring of surface proteins, we analyzed variants of staphylococcal protein A, an immunoglobulin binding protein with an LPXTG sorting signal. Deletion of the YSIR sequence or replacement of G or S significantly reduced the rate of signal peptide processing of protein A precursors. In contrast, cell wall anchoring or the functional display of protein A was not affected. The fusion of cell wall sorting signals to reporter proteins bearing N-terminal signal peptides with or without the YSIRK-G/S motif resulted in hybrid proteins that were anchored in a manner similar to that of wild-type protein A. The requirement of the YSIRK-G/S motif for efficient secretion implies the existence of a specialized mode of substrate recognition by the secretion pathway of gram-positive cocci. It seems, however, that this mechanism is not essential for surface protein anchoring to the cell wall envelope.
Figures
References
-
- Benson, S. A., M. N. Hall, and T. J. Silhavy. 1985. Genetic analysis of protein export in Escherichia coli K12. Annu. Rev. Biochem. 54:101-134. - PubMed
-
- Bernstein, H. D., M. A. Poritz, K. Strub, P. J. Hoben, S. Brenner, and P. Walter. 1989. Model for signal sequence recognition from amino-acid sequence of 54K subunit of signal recognition particle. Nature 340:482-486. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
