

Haplotypic analysis of the TNF locus by association efficiency and entropy
- PMID: 12702205
- PMCID: PMC154575
- DOI: 10.1186/gb-2003-4-4-r24
Haplotypic analysis of the TNF locus by association efficiency and entropy
Abstract
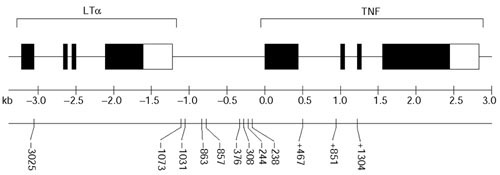
Background: To understand the causal basis of TNF associations with disease, it is necessary to understand the haplotypic structure of this locus. We genotyped 12 single-nucleotide polymorphisms (SNPs) distributed over 4.3 kilobases in 296 healthy, unrelated Gambian and Malawian adults. We generated 592 high-quality haplotypes by integrating family- and population-based reconstruction methods.
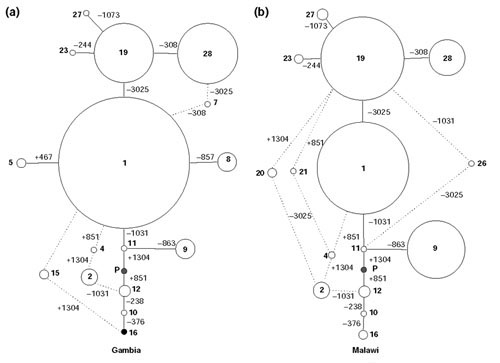
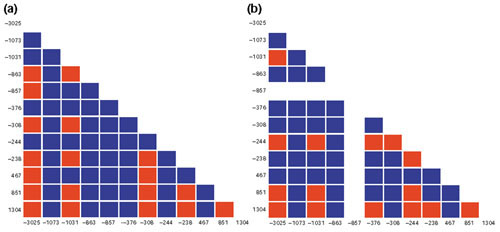
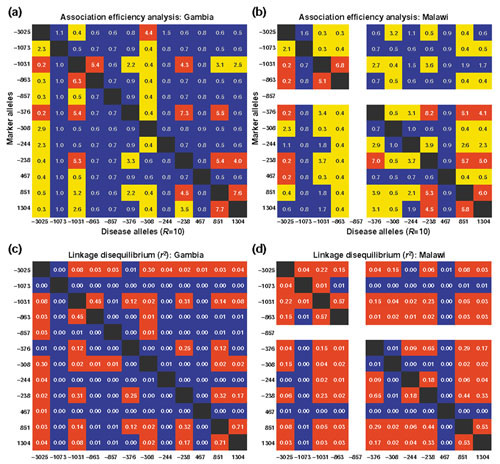
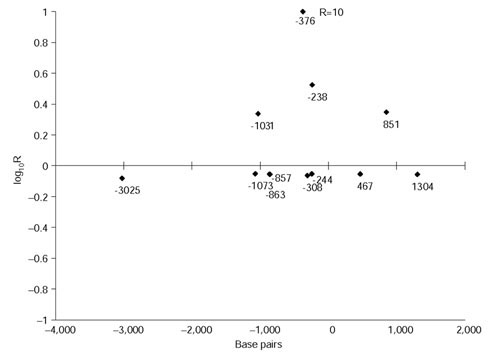
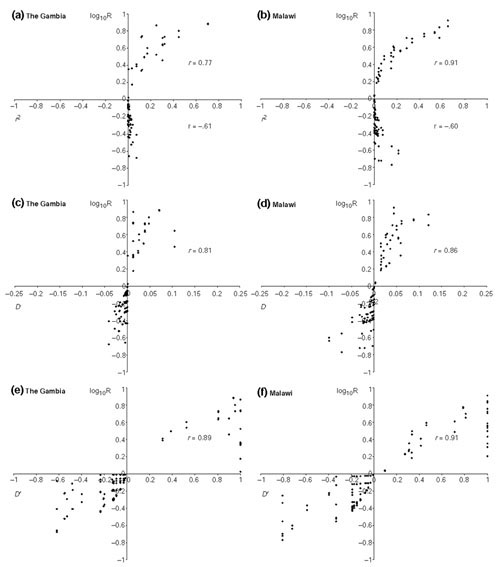
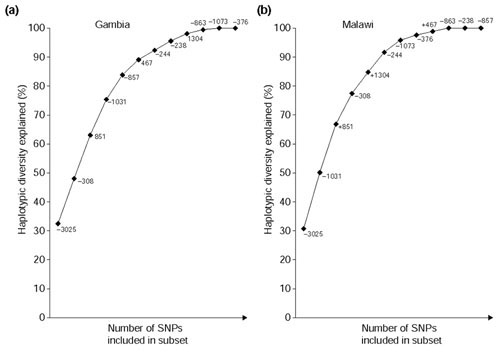
Results: We found 32 different haplotypes, of which 13 were shared between the two populations. Both populations were haplotypically diverse (gene diversity = 0.80, Gambia; 0.85, Malawi) and significantly differentiated (p < 10-5 by exact test). More than a quarter of marker pairs showed evidence of intragenic recombination (29% Gambia; 27% Malawi). We applied two new methods of analyzing haplotypic data: association efficiency analysis (AEA), which describes the ability of each SNP to detect every other SNP in a case-control scenario; and the entropy maximization method (EMM), which selects the subset of SNPs that most effectively dissects the underlying haplotypic structure. AEA revealed that many SNPs in TNF are poor markers of each other. The EMM showed that 8 of 12 SNPs (Gambia) and 7 of 12 SNPs (Malawi) are required to describe 95% of the haplotypic diversity.
Conclusions: The TNF locus in the Gambian and Malawi sample is haplotypically diverse and has a rich history of intragenic recombination. As a consequence, a large proportion of TNF SNPs must be typed to detect a disease-modifying SNP at this locus. The most informative subset of SNPs to genotype differs between the two populations.
Figures
References
-
- McGuire W, Hill AV, Allsopp CE, Greenwood BM, Kwiatkowski D. Variation in the TNF-alpha promoter region associated with susceptibility to cerebral malaria. Nature. 1994;371:508–510. - PubMed
-
- McGuire W, Knight JC, Hill AV, Allsopp CE, Greenwood BM, Kwiatkowski D. Severe malarial anemia and cerebral malaria are associated with different tumor necrosis factor promoter alleles. J Infect Dis. 1999;179:287–290. - PubMed
-
- Knight JC, Udalova I, Hill AV, Greenwood BM, Peshu N, Marsh K, Kwiatkowski D. A polymorphism that affects OCT-1 binding to the TNF promoter region is associated with severe malaria. Nat Genet. 1999;22:145–150. - PubMed
-
- Knight JC, Kwiatkowski D. Inherited variability of tumor necrosis factor production and susceptibility to infectious disease. Proc Assoc Am Physicians. 1999;111:290–298. - PubMed
-
- Dunstan SJ, Stephens HA, Blackwell JM, Duc CM, Lanh MN, Dudbridge F, Phuong CX, Luxemburger C, Wain J, Ho VA, et al. Genes of the class II and class III major histocompatibility complex are associated with typhoid fever in Vietnam. J Infect Dis. 2001;183:261–268. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
