

doi: 10.1073/pnas.0531307100.
Epub 2003 Apr 17.
The riboswitch-mediated control of sulfur metabolism in bacteria
Affiliations
- PMID: 12702767
- PMCID: PMC154296
- DOI: 10.1073/pnas.0531307100
Item in Clipboard
The riboswitch-mediated control of sulfur metabolism in bacteria
Proc Natl Acad Sci U S A.
.
Abstract
Many operons in Gram-positive bacteria that are involved in methionine (Met) and cysteine (Cys) biosynthesis possess an evolutionarily conserved regulatory leader sequence (S-box) that positively controls these genes in response to methionine starvation. Here, we demonstrate that a feed-back regulation mechanism utilizes S-adenosyl-methionine as an effector. S-adenosyl-methionine directly and specifically binds to the nascent S-box RNA, causing an intrinsic terminator to form and interrupt transcription prematurely. The S-box leader RNA thus expands the family of newly discovered riboswitches, i.e., natural regulatory RNA aptamers that seem to sense small molecules ranging from amino acid derivatives to vitamins.
Figures
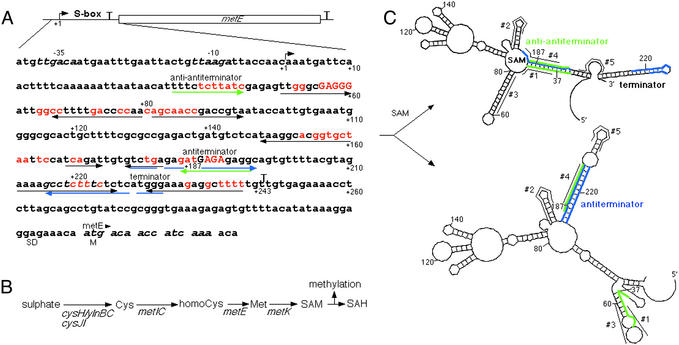
Model for the SAM-mediated attenuation control of the S-box regulon. (A) S-box leader region of the metE gene. Numbers indicate positions beginning at the +1 start of transcription. The S-box is defined as a region that includes the evolutionarily conserved bases shown in red (11). Mutations that were analyzed in detail in this work are capitalized. Blue arrows show the antiterminator and green arrows the anti-antiterminator. Black arrows show potential stem-loops including that of the terminator. The termination points are indicated by “T.” SD, Shine-Dalgarno sequence; M, the first Met of metE gene. (B) The pathway of sulfate assimilation and Met and SAM biosynthesis. Only S-box family genes that are repressed by SAM are indicated (11, 15). The function of these genes is as follows: ylnB, ATP sulfurylase; ylnC, adenosine phosphosulfate kinase; cysH, 3′-phosphoadenosine 5′-phosphosulfate sulfotransferase; cysJI, NADH-sulfite reductase; metI, cystathionine γ-synthase/O-acetylhomoserine sulfhydrylase; metC, cystathionine β-lyase; metK, SAM synthetase. Various methyl transferases convert SAM to SAH during methylation. (C) SAM-directed terminator/antiterminator riboswitch. Two alternative structures of S-box, the anti-antiterminator (Upper) and antiterminator (Lower), were calculated by using the Zuker–Turner algorithm of free energy minimization (18). The antiterminator sequence is in blue and anti-antiterminator in green. The program predicts the anti-antiterminator structure by default and the antitermination structure by forcing the complementary “blue” bases to be paired. According to the model, the anti-antiterminator structure folds only in the presence of SAM. Numbers indicate the positions in the RNA in relation to the 5′ terminus (+1). These reference numbers are also shown in (A). #1–#5, the antisense DNA oligos that were used to probe the structure of S-box in the experiment of Fig. 3. The oligos are shown next to their annealing targets.
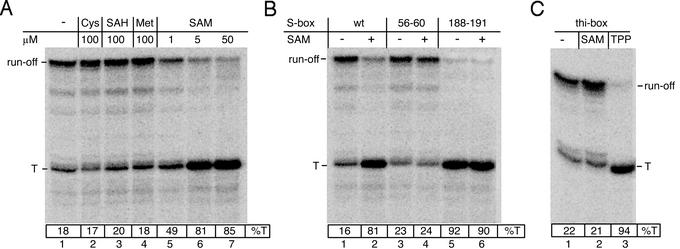
SAM-mediated transcription termination in vitro. (A) Effect of small molecules on termination by B. subtilis RNAP. The autoradiogram of the 6% sequencing PAGE shows 32P-labeled full size (run-off) and terminated (T) RNA products from a reconstituted single round transcription reaction (see Experimental Procedures). T, terminated transcript; SAM, Met, Cys, and SAH were added to the final concentrations indicated at the top. (B) Effect of S-box mutations on termination. Mutations are as follows: “56–60,” G56A, A57T, G58C, G59A, G60C; and “188–191” (the antiterminator), G188C, A189T, G190A, and A191T. The concentration of SAM was 5 μM. (C) Effect of SAM and TPP on termination of the thi-box leader transcript. The autoradiogram of the 6% sequencing PAGE shows 32P-labeled full size (run-off) and terminated (T) RNA products from the reconstituted single round transcription reaction (see Experimental Procedures). TPP and SAM were added to 50 μM.
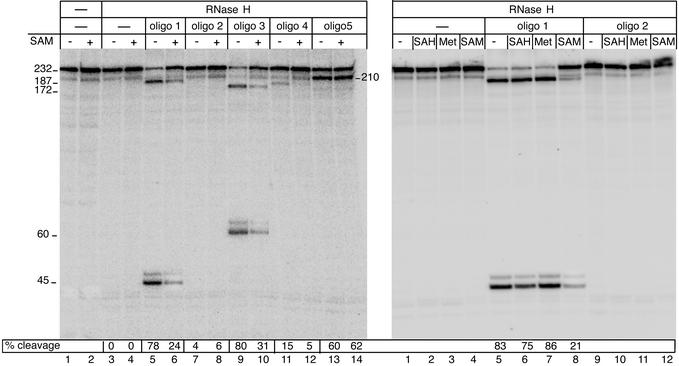
The effect of SAM on the structure of the pure S-box RNA. RNA structural transition is monitored by using the antisense DNA oligonucleotides and RNase H (5). The truncated S-box-leader template lacking the terminator was used to obtain the 232-nt transcript. Its 5′ proximal region was 32P-labeled (see Experimental Procedures). The transcript was extracted from the transcription reaction with phenol/chloroform, followed by ethanol precipitation. The pure RNA was dissolved in TB with or without SAM. The mixture was then heated to 80°C for 5 min and slowly cooled to room temperature. The changes in RNA structure during refolding in the presence of SAM (5 μM) were monitored by annealing oligos 1–5, followed by RNase H cleavage. As the radiogram shows, the sensitivity of refolded RNA to RNase H was greatly suppressed by SAM (Left, lanes 6 and 8) but not by Met (100 μM) or SAH (100 μM; Right, lanes 6 and 7). This experiment indicates that the formation of a specific binary complex between the S-box RNA and SAM that triggers the conformational change in the leader RNA occurs during S-box RNA folding, independently of other components of the elongation complex. Numbers on the right and left indicate the size of the RNase H cleavage products. The annealing sites for oligos are shown in Fig. 1C.
References
-
- Henkin T M, Yanofsky C. BioEssays. 2002;24:700–707. - PubMed
-
- Nahvi A, Sudarsan N, Ebert M S, Zou X, Brown K L, Breaker R R. Chem Biol. 2002;9:1043–1049. - PubMed
-
- Winkler W, Nahvi A, Breaker R R. Nature. 2002;419:952–956. - PubMed
-
- Mironov A S, Gusarov I, Rafikov R, Lopez L E, Shatalin K, Kreneva R A, Perumov D A, Nudler E. Cell. 2002;111:747–756. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
