

Otubains: a new family of cysteine proteases in the ubiquitin pathway
- PMID: 12704427
- PMCID: PMC1319179
- DOI: 10.1038/sj.embor.embor824
Otubains: a new family of cysteine proteases in the ubiquitin pathway
Abstract
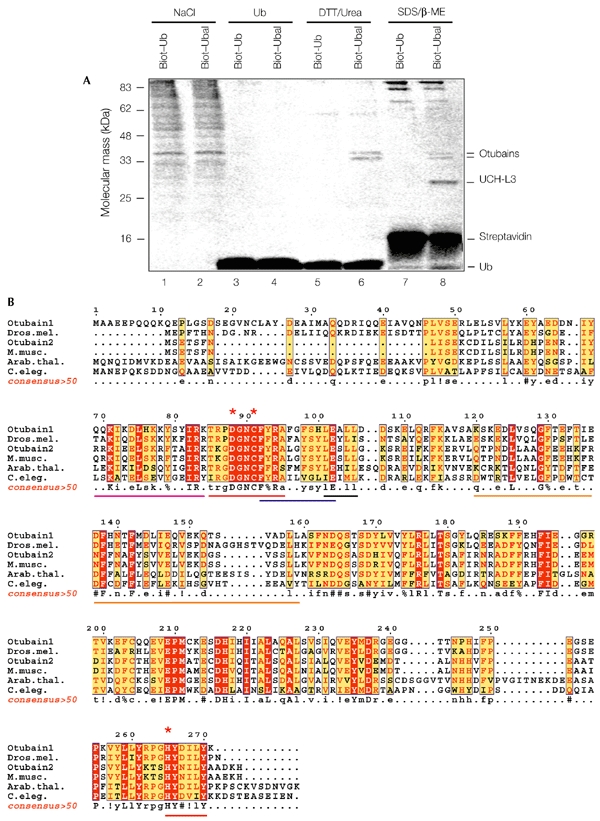
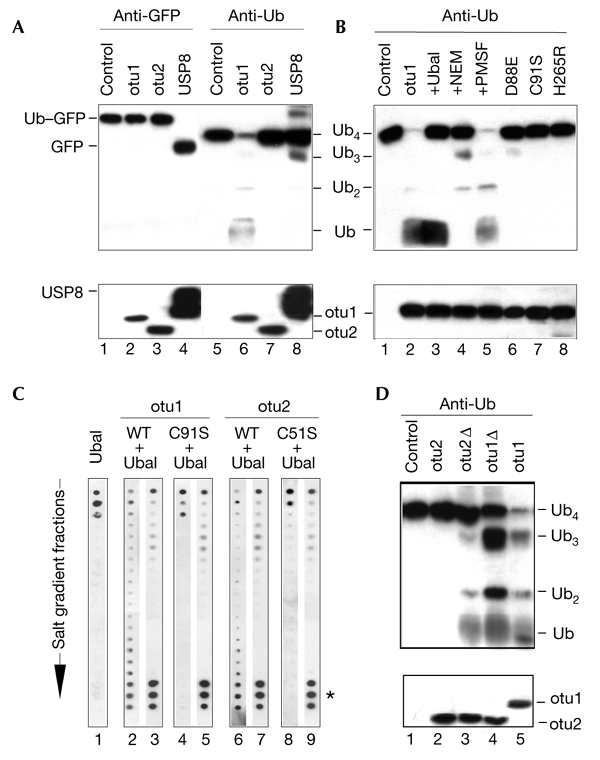
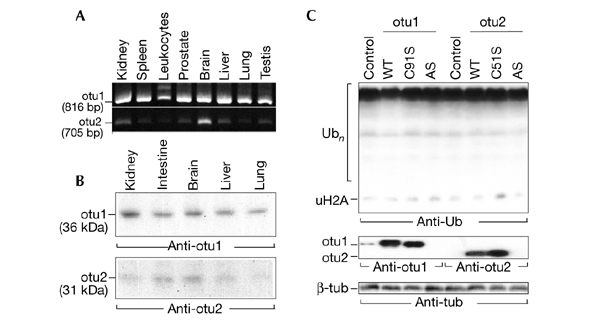
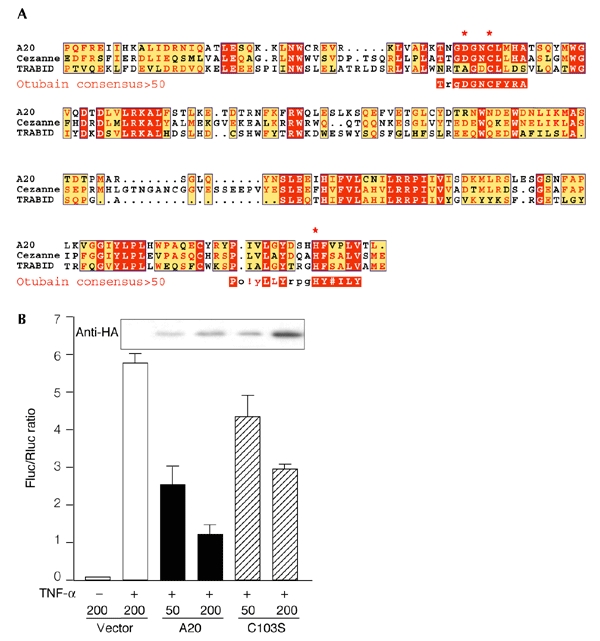
The modification of cellular proteins by ubiquitin (Ub) is an important event that underlies protein stability and function in eukaryotes. Protein ubiquitylation is a dynamic and reversible process; attached Ub can be removed by deubiquitylating enzymes (DUBs), a heterogeneous group of cysteine proteases that cleave proteins precisely at the Ub-protein bond. Two families of DUBs have been identified previously. Here, we describe new, highly specific Ub iso-peptidases, that have no sequence homology to known DUBs, but which belong to the OTU (ovarian tumour) superfamily of proteins. Two novel proteins were isolated from HeLa cells by affinity purification using the DUB-specific inhibitor, Ub aldehyde (Ubal). We have named these proteins otubain 1 and otubain 2, for OTU-domain Ubal-binding protein. Functional analysis of otubains shows that the OTU domain contains an active cysteine protease site.
Figures
References
-
- Beyaert R., Heyninck K. & Van Huffel S. (2000) A20 and A20-binding proteins as cellular inhibitors of nuclear factor-κB-dependent gene expression and apoptosis. Biochem. Pharmacol., 60, 1143–1151. - PubMed
-
- Borodovsky A., Ovaa H., Kolli N., Gan-Erdene T., Wilkinson K.D., Ploegh H.L. & Kessler B.M. (2002) Chemistry-based functional proteomics reveals novel members of the deubiquitinating enzyme family. Chem. Biol., 9, 1149–1159. - PubMed
-
- D'Andrea A. & Pellman D. (1998) Deubiquitinating enzymes: a new class of biological regulators. Crit. Rev. Biochem. Mol. Biol., 33, 337–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
