

The DNA methyltransferases associate with HP1 and the SUV39H1 histone methyltransferase
- PMID: 12711675
- PMCID: PMC154218
- DOI: 10.1093/nar/gkg332
The DNA methyltransferases associate with HP1 and the SUV39H1 histone methyltransferase
Abstract
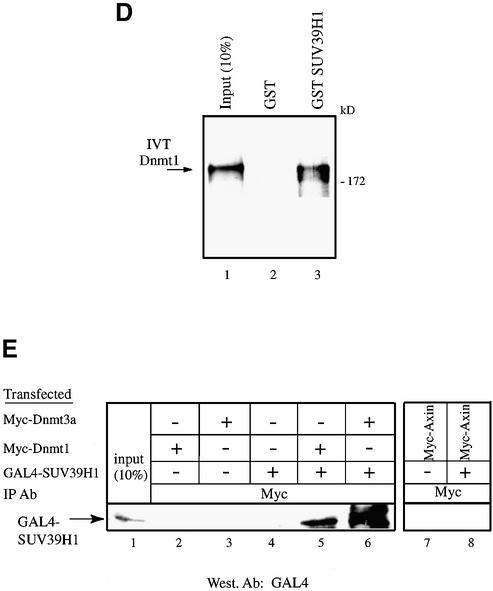
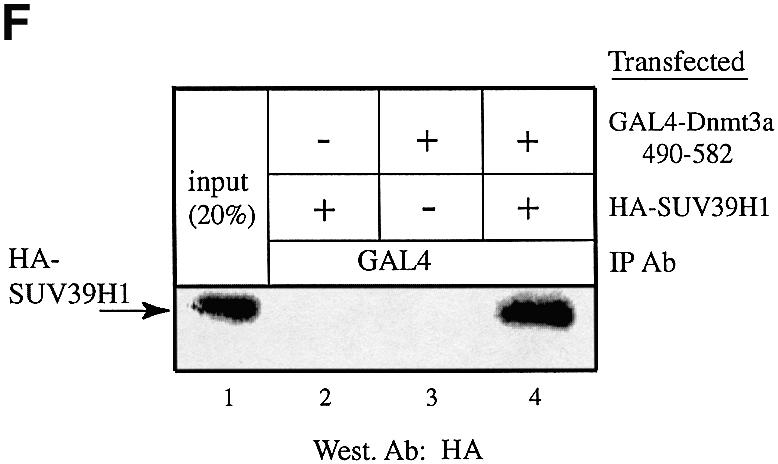
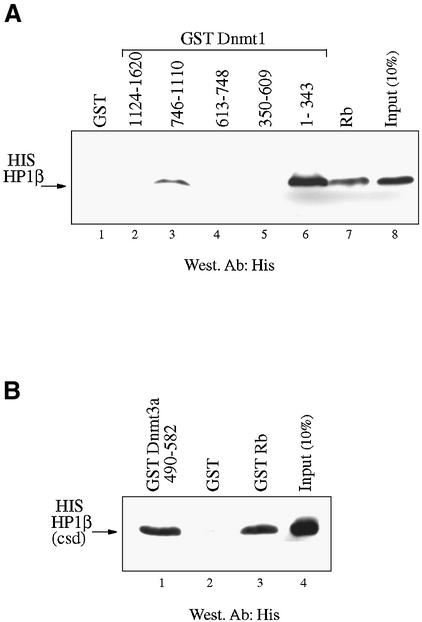
The DNA methyltransferases, Dnmts, are the enzymes responsible for methylating DNA in mammals, which leads to gene silencing. Repression by DNA methylation is mediated partly by recruitment of the methyl-CpG-binding protein MeCP2. Recently, MeCP2 was shown to associate and facilitate histone methylation at Lys9 of H3, which is a key epigenetic modification involved in gene silencing. Here, we show that endogenous Dnmt3a associates primarily with histone H3-K9 methyltransferase activity as well as, to a lesser extent, with H3-K4 enzymatic activity. The association with enzymatic activity is mediated by the conserved PHD-like motif of Dnmt3a. The H3-K9 histone methyltransferase that binds Dnmt3a is likely the H3-K9 specific SUV39H1 enzyme since we find that it interacts both in vitro and in vivo with Dnmt3a, using its PHD-like motif. We find that SUV39H1 also binds to Dnmt1 and, consistent with these interactions, SUV39H1 can purify DNA methyltransferase activity from nuclear extracts. In addition, we show that HP1beta, a SUV39H1-interacting partner, binds directly to Dnmt1 and Dnmt3a and that native HP1beta associates with DNA methyltransferase activity. Our data show a direct connection between the enzymes responsible for DNA methylation and histone methylation. These results further substantiate the notion of a self-reinforcing repressive chromatin state through the interplay between these two global epigenetic modifications.
Figures

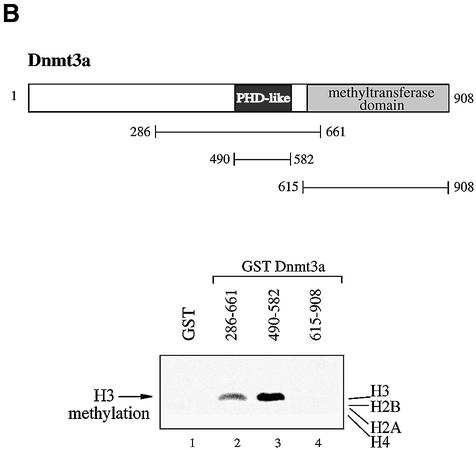
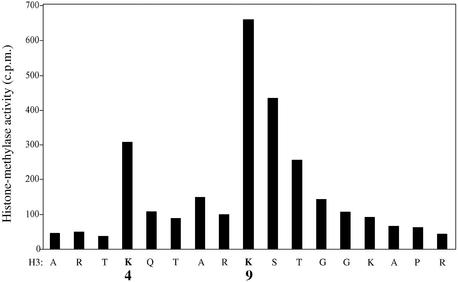
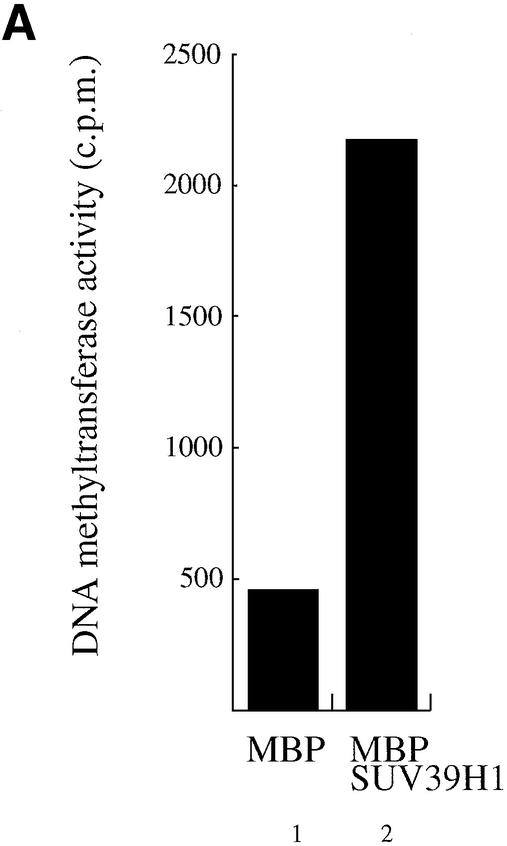
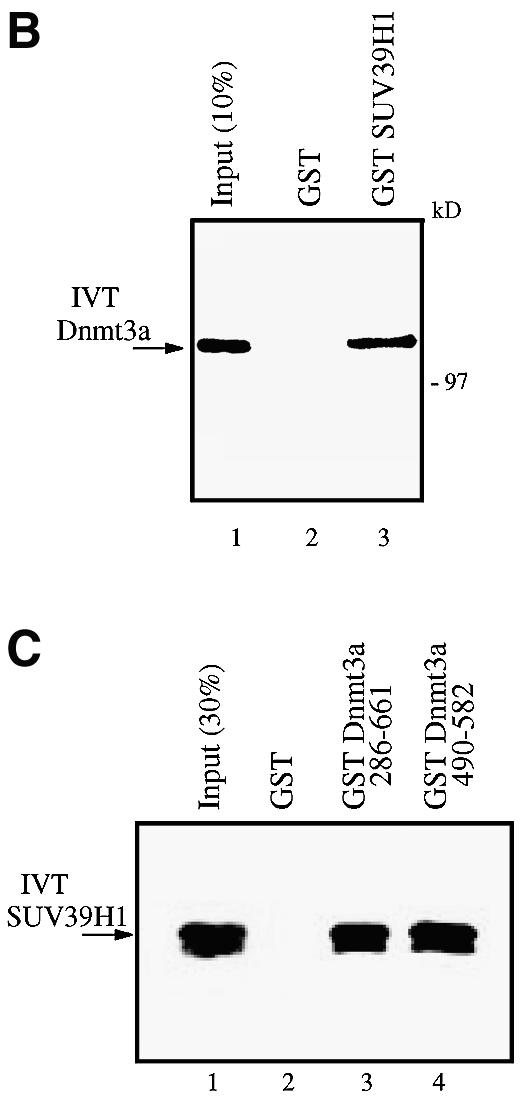


References
-
- Bird A. (2002) DNA methylation patterns and epigenetic memory. Genes Dev., 16, 6–21. - PubMed
-
- Bestor T.H. (2000) The DNA methyltransferases of mammals. Hum. Mol. Genet., 9, 2395–2402. - PubMed
-
- Li E., Bestor,T.H. and Jaenisch,R. (1992) Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell, 69, 915–926. - PubMed
-
- Okano M., Bell,D.W., Haber,D.A. and Li,E. (1999) DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell, 99, 247–257. - PubMed
-
- Bird A.P. and Wolffe,A.P. (1999) Methylation-induced repression—belts, braces and chromatin. Cell, 99, 451–454. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
