

Circadian phase-dependent modulation of cGMP-gated channels of cone photoreceptors by dopamine and D2 agonist
- PMID: 12716922
- PMCID: PMC2667621
- DOI: 10.1523/JNEUROSCI.23-08-03145.2003
Circadian phase-dependent modulation of cGMP-gated channels of cone photoreceptors by dopamine and D2 agonist
Abstract
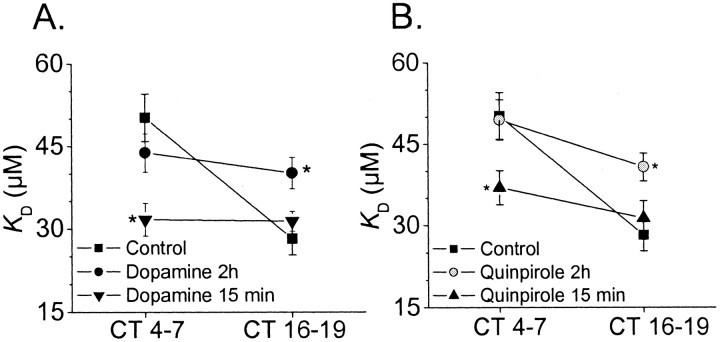
The affinity of cGMP-gated ion channels (CNGCs) for cGMP in chick retinal cone photoreceptors is under circadian control. Here we report that dopamine (DA) and D2 receptor agonists evoke phase-dependent shifts in the affinity of CNGCs for activating ligand. Inside-out patch recordings from cultured chick cones were performed at circadian time (CT) 4-7 and CT 16-19 on the second day of constant darkness. Exposing intact cells to DA or the D2 agonist quinpirole for 2 hr before patch excision caused a significant increase in the K(D) for cGMP during the night (CT 16-19) but had no effect during the day (CT 4-7). DA or quinpirole treatment had no effect on the Hill slope or the average number of channels per patch. The effect of DA was blocked by the D2 antagonist eticlopride and was not mimicked by D1 agonists or blocked by D1 antagonists. By contrast, a brief (15 min) exposure to DA or quinpirole caused a decrease in K(D) during the subjective day and had no effect during the subjective night. Thus, the effect of D2 agonists depends on both the duration of agonist exposure and the time of day. Application of DA or quinpirole evoked a transient activation of the MAP kinase Erk (extracellular signal-related kinase) during the day but caused a sustained inhibition during the night. Conversely, D2 agonists caused activation of Ca2+/calmodulin-dependent protein kinase II during the night and inhibited this enzyme during the day. A circadian oscillator in cones appears to regulate the nature of the transduction cascade used by D2 receptors.
Figures







Similar articles
-
Circadian regulation of cGMP-gated cationic channels of chick retinal cones. Erk MAP Kinase and Ca2+/calmodulin-dependent protein kinase II.Neuron. 2001 Jan;29(1):255-66. doi: 10.1016/s0896-6273(01)00195-7. Neuron. 2001. PMID: 11182096
-
Tyrosine phosphorylation of cGMP-gated ion channels is under circadian control in chick retina photoreceptors.Invest Ophthalmol Vis Sci. 2007 Feb;48(2):901-6. doi: 10.1167/iovs.06-0824. Invest Ophthalmol Vis Sci. 2007. PMID: 17251493 Free PMC article.
-
Circadian regulation of cGMP-gated channels of vertebrate cone photoreceptors: role of cAMP and Ras.J Neurosci. 2004 Feb 11;24(6):1296-304. doi: 10.1523/JNEUROSCI.3560-03.2004. J Neurosci. 2004. PMID: 14960600 Free PMC article.
-
Tuning outer segment Ca2+ homeostasis to phototransduction in rods and cones.Adv Exp Med Biol. 2002;514:179-203. doi: 10.1007/978-1-4615-0121-3_11. Adv Exp Med Biol. 2002. PMID: 12596922 Review.
-
Circadian regulation of ion channels and their functions.J Neurochem. 2009 Aug;110(4):1150-69. doi: 10.1111/j.1471-4159.2009.06223.x. Epub 2009 Jun 15. J Neurochem. 2009. PMID: 19549279 Free PMC article. Review.
Cited by
-
Loss of cone cyclic nucleotide-gated channel leads to alterations in light response modulating system and cellular stress response pathways: a gene expression profiling study.Hum Mol Genet. 2013 Oct 1;22(19):3906-19. doi: 10.1093/hmg/ddt245. Epub 2013 Jun 4. Hum Mol Genet. 2013. PMID: 23740940 Free PMC article.
-
Rhythmic expression of microRNA-26a regulates the L-type voltage-gated calcium channel alpha1C subunit in chicken cone photoreceptors.J Biol Chem. 2009 Sep 18;284(38):25791-803. doi: 10.1074/jbc.M109.033993. Epub 2009 Jul 16. J Biol Chem. 2009. PMID: 19608742 Free PMC article.
-
The Contribution of L-Type Cav1.3 Channels to Retinal Light Responses.Front Mol Neurosci. 2017 Dec 5;10:394. doi: 10.3389/fnmol.2017.00394. eCollection 2017. Front Mol Neurosci. 2017. PMID: 29259539 Free PMC article.
-
Modulation of the ERK1/2-MMP-2 pathway in the sclera of guinea pigs following induction of myopia by flickering light.Exp Ther Med. 2021 Apr;21(4):371. doi: 10.3892/etm.2021.9802. Epub 2021 Feb 19. Exp Ther Med. 2021. PMID: 33732344 Free PMC article.
-
Circadian regulation in the retina: From molecules to network.Eur J Neurosci. 2020 Jan;51(1):194-216. doi: 10.1111/ejn.14185. Epub 2018 Oct 24. Eur J Neurosci. 2020. PMID: 30270466 Free PMC article. Review.
References
-
- Adachi A, Nogi T, Ebihara S. Phase-relationship and mutual effects between circadian rhythms of ocular melatonin and dopamine in the pigeon. Brain Res. 1998;792:361–369. - PubMed
-
- Adams JP, Anderson AE, Varga AW, Dineley KT, Cook RG, Pfaffinger PJ, Sweatt JD. The A-type potassium channel Kv4.2 is a substrate for the mitogen-activated protein kinase ERK. J Neurochem. 2000;75:2277–2287. - PubMed
-
- Adler R, Hatlee M. Plasticity and differentiation of embryonic retinal cells after terminal mitosis. Science. 1989;243:391–393. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous