

Mice lacking the giant protocadherin mFAT1 exhibit renal slit junction abnormalities and a partially penetrant cyclopia and anophthalmia phenotype
- PMID: 12724416
- PMCID: PMC164754
- DOI: 10.1128/MCB.23.10.3575-3582.2003
Mice lacking the giant protocadherin mFAT1 exhibit renal slit junction abnormalities and a partially penetrant cyclopia and anophthalmia phenotype
Abstract
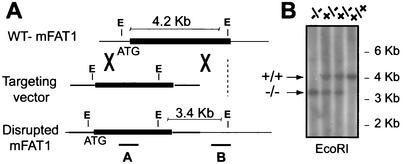
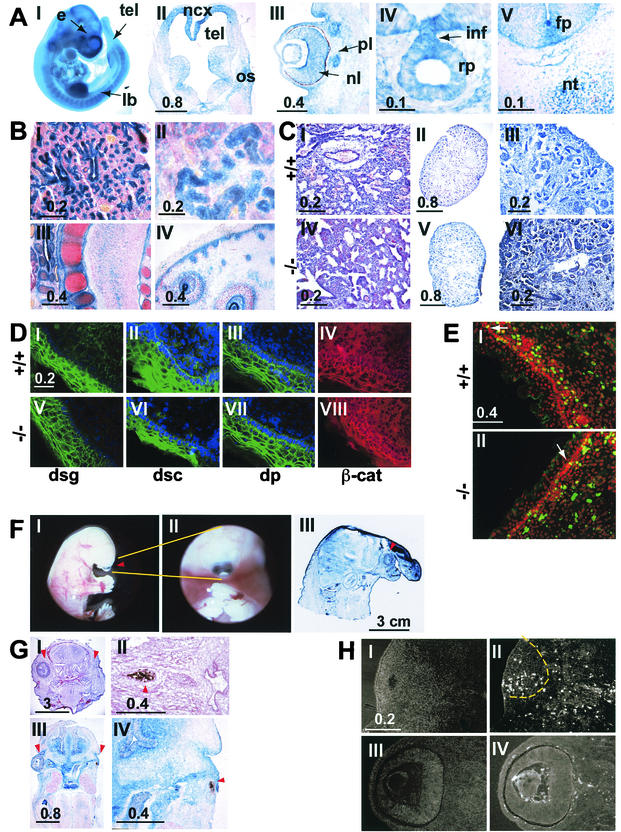
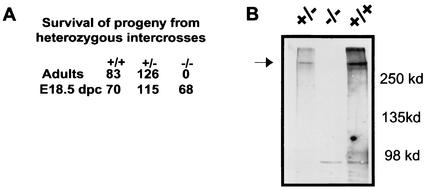
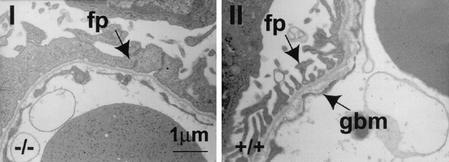
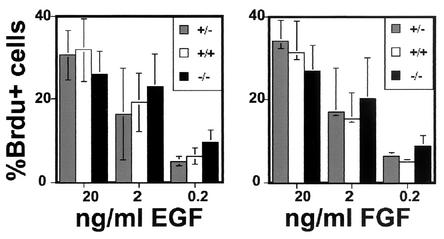
While roles in adhesion and morphogenesis have been documented for classical cadherins, the nonclassical cadherins are much less well understood. Here we have examined the functions of the giant protocadherin FAT by generating a transgenic mouse lacking mFAT1. These mice exhibit perinatal lethality, most probably caused by loss of the renal glomerular slit junctions and fusion of glomerular epithelial cell processes (podocytes). In addition, some mFAT1(-/-) mice show defects in forebrain development (holoprosencephaly) and failure of eye development (anophthalmia). In contrast to Drosophila, where FAT acts as a tumor suppressor gene, we found no evidence for abnormalities of proliferation in two tissues (skin and central nervous system [CNS]) containing stem and precursor cell populations and in which FAT is expressed strongly. Our results confirm a necessary role for FAT1 in the modified adhesion junctions of the renal glomerular epithelial cell and reveal hitherto unsuspected roles for FAT1 in CNS development.
Figures
References
-
- Adler, R., and T. L. Belecky-Adams. 2002. The role of bone morphogenetic proteins in the differentiation of the ventral optic cup. Development 129:3161-3171. - PubMed
-
- Alagramam, K. N., H. Yuan, M. H. Kuehn, C. L. Murcia, S. Wayne, C. R. Srisailpathy, R. B. Lowry, R. Knaus, L. Van Laer, F. P. Bernier, S. Schwartz, C. Lee, C. C. Morton, R. F. Mullins, A. Ramesh, G. Van Camp, G. S. Hageman, R. P. Woychik, R. J. Smith, and G. S. Hagemen. 2001. Mutations in the novel protocadherin PCDH15 cause Usher syndrome type 1F. Hum. Mol. Genet. 10:1709-1718. - PubMed
-
- Bradley, R. S., A. Espeseth, and C. Kintner. 1998. NF-protocadherin, a novel member of the cadherin superfamily, is required for Xenopus ectodermal differentiation. Curr. Biol. 8:325-334. - PubMed
-
- Chiang, C., Y. Litingtung, E. Lee, K. E. Young, J. L. Corden, H. Westphal, and P. A. Beachy. 1996. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 383:407-413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases