

Modulation of mitochondrial function by endogenous Zn2+ pools
- PMID: 12724524
- PMCID: PMC156342
- DOI: 10.1073/pnas.1031598100
Modulation of mitochondrial function by endogenous Zn2+ pools
Abstract
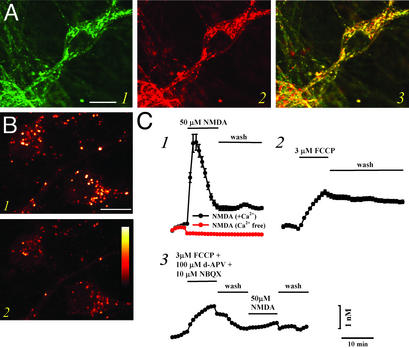
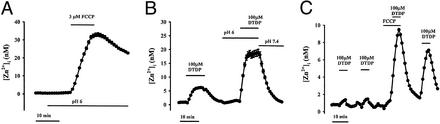
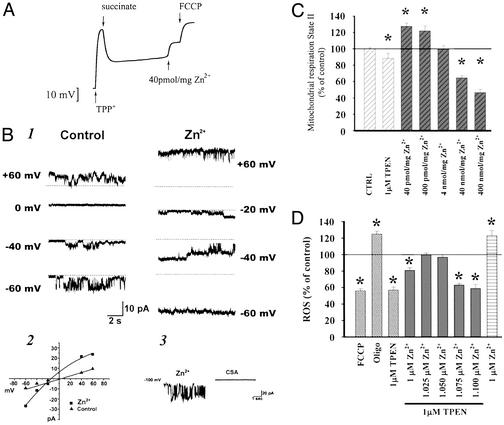
Recent evidence suggests that intracellular Zn(2+) accumulation contributes to the neuronal injury that occurs in epilepsy or ischemia in certain brain regions, including hippocampus, amygdala, and cortex. Although most attention has been given to the vesicular Zn(2+) that is released into the synaptic space and may gain entry to postsynaptic neurons, recent studies have highlighted pools of intracellular Zn(2+) that are mobilized in response to stimulation. One such Zn(2+) pool is likely bound to cytosolic proteins, like metallothioneins. Applying imaging techniques to cultured cortical neurons, this study provides novel evidence for the presence of a mitochondrial pool distinct from the cytosolic protein or ligand-bound pool. These pools can be pharmacologically mobilized largely independently of each other, with Zn(2+) release from one resulting in apparent net Zn(2+) transfer to the other. Further studies found evidence for complex and potent effects of Zn(2+) on isolated brain mitochondria. Submicromolar levels, comparable to those that might occur on strong mobilization of intracellular compartments, induced membrane depolarization (loss of Deltapsi(m)), increases in currents across the mitochondrial inner membrane as detected by direct patch clamp recording of mitoplasts, increased O(2) consumption and decreased reactive oxygen species (ROS) generation, whereas higher levels decreased O(2) consumption and increased ROS generation. Finally, strong mobilization of protein-bound Zn(2+) appeared to induce partial loss of Deltapsi(m), suggesting that movement of Zn(2+) between cytosolic and mitochondrial pools might be of functional significance in intact neurons.
Figures

References
-
- Frederickson C J, Bush A I. Biometals. 2001;14:353–366. - PubMed
-
- Aschner M, Cherian M G, Klaassen C D, Palmiter R D, Erickson J C, Bush A I. Toxicol Appl Pharmacol. 1997;142:229–242. - PubMed
-
- Kagi J H R. In: Metallothionein III: Biological Roles and Medical Implications. Kimura M, editor. Basel: Birkhauser; 1993. pp. 29–55.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
