

Dominant-negative receptor uncovers redundancy in the Arabidopsis ERECTA Leucine-rich repeat receptor-like kinase signaling pathway that regulates organ shape
- PMID: 12724536
- PMCID: PMC153719
- DOI: 10.1105/tpc.010413
Dominant-negative receptor uncovers redundancy in the Arabidopsis ERECTA Leucine-rich repeat receptor-like kinase signaling pathway that regulates organ shape
Abstract
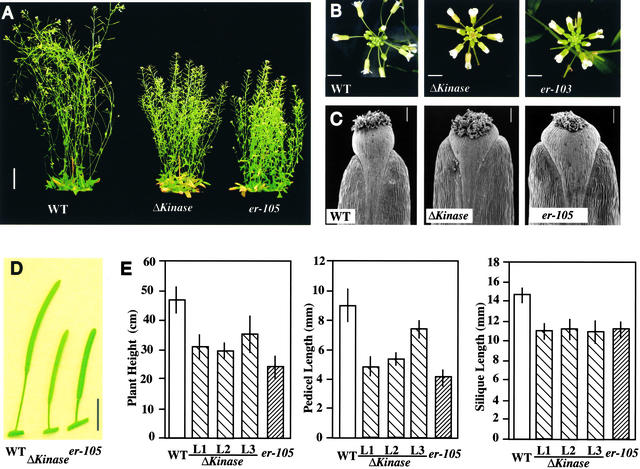
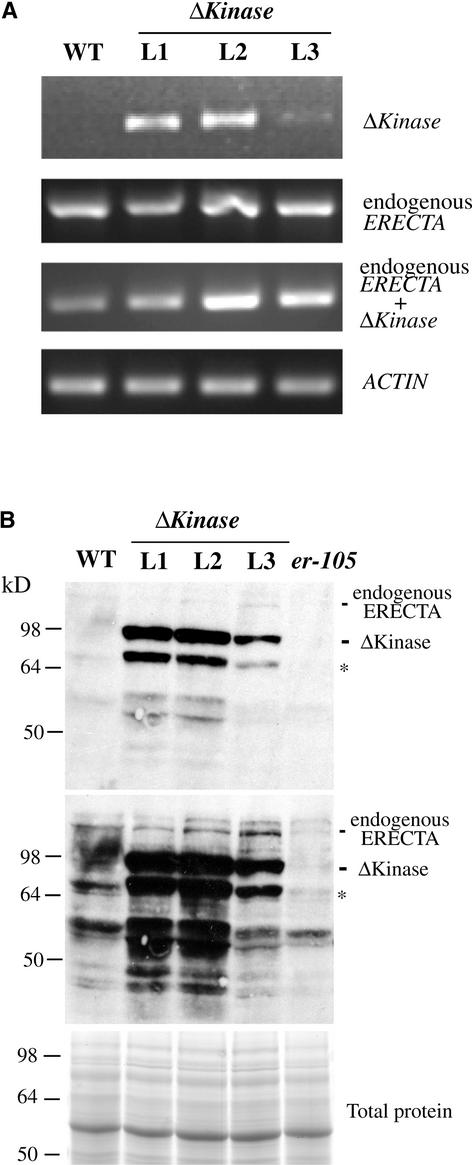
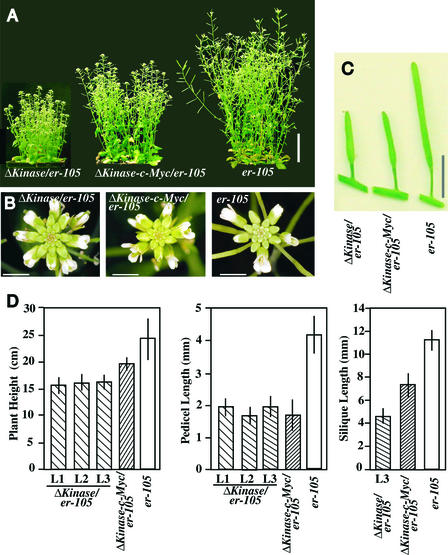
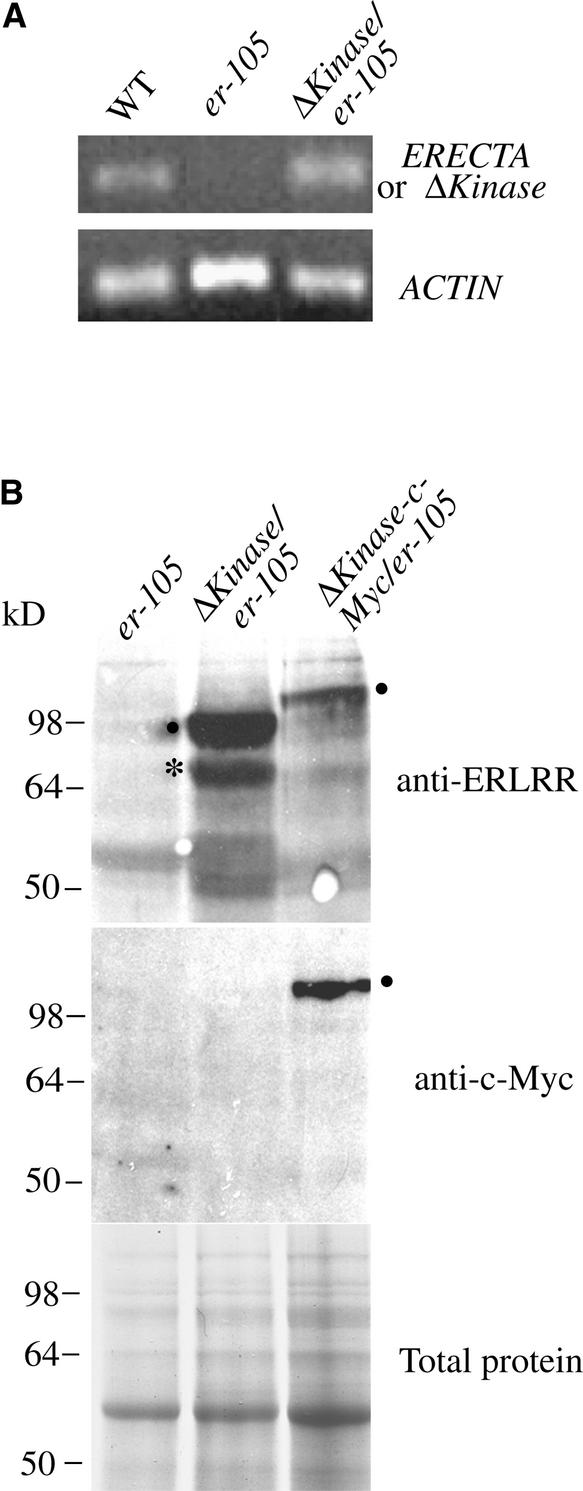
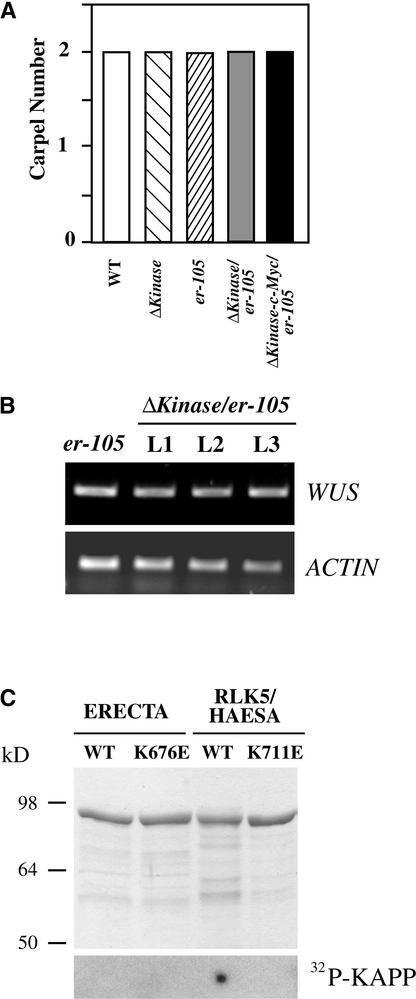
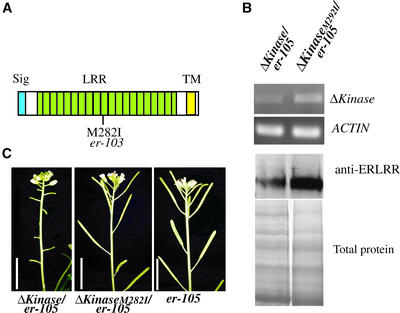
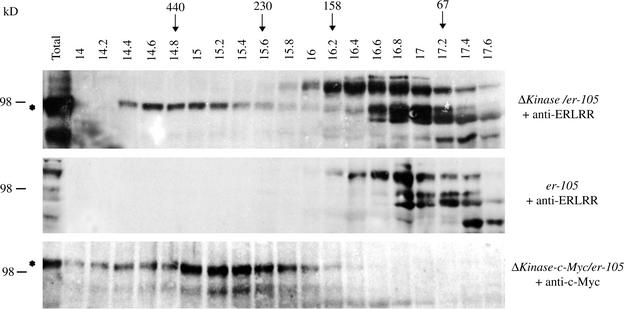
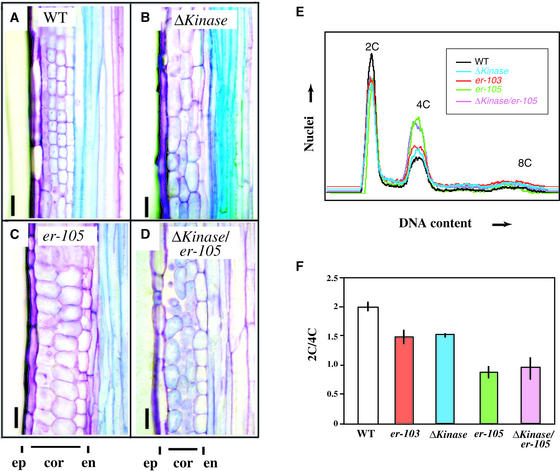
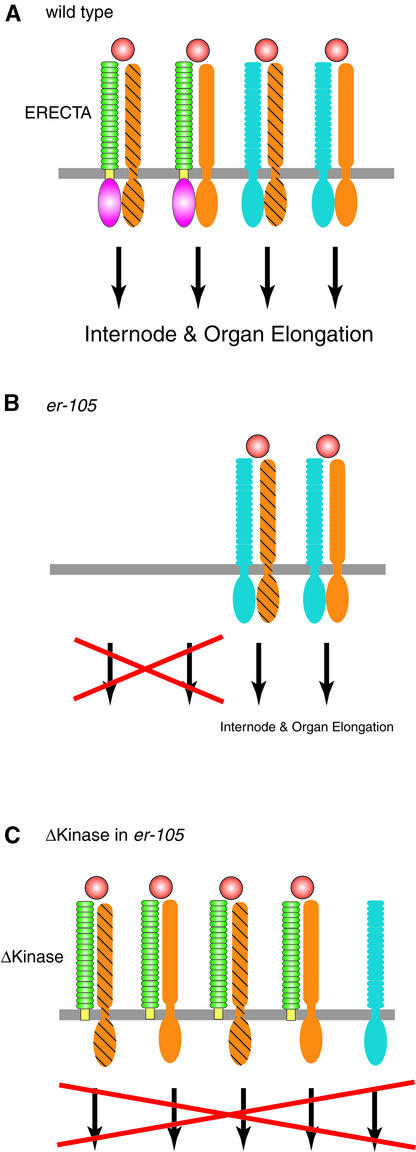
Arabidopsis ERECTA, a Leu-rich repeat receptor-like Ser/Thr kinase (LRR-RLK), regulates organ shape and inflorescence architecture. Here, we show that a truncated ERECTA protein that lacks the cytoplasmic kinase domain (DeltaKinase) confers dominant-negative effects when expressed under the control of the native ERECTA promoter and terminator. Transgenic plants expressing DeltaKinase displayed phenotypes, including compact inflorescence and short, blunt siliques, that are characteristic of loss-of-function erecta mutant plants. The DeltaKinase fragment migrated as a stable approximately 400-kD protein complex in the complete absence of the endogenous ERECTA protein and significantly exaggerated the growth defects of the null erecta plants. A functional LRR domain of DeltaKinase was required for dominant-negative effects. Accumulation of DeltaKinase did not interfere with another LRR-RLK signaling pathway (CLAVATA1), which operates in the same cells as ERECTA but has a distinct biological function. Both the erecta mutation and DeltaKinase expression conferred a lesser number of large, disorganized, and expanded cortex cells, which are associated with an increased level of somatic endoploidy. These findings suggest that functionally redundant RLK signaling pathways, including ERECTA, are required to fine-tune the proliferation and growth of cells in the same tissue type during Arabidopsis organogenesis.
Figures
References
-
- Amaya, E., Musci, T., and Kirschner, M. (1991). Expression of a dominant-negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell 66, 257–270. - PubMed
-
- Ataliotis, P., and Mercola, M. (1997). Distribution and function of platelet-derived growth factors and their receptors during embryogenesis. Int. Rev. Cytol. 172, 95–125. - PubMed
-
- Bechtold, N., Ellis, J.G., and Pelletier, G. (1993). In planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C. R. Acad. Sci. Paris 316, 1194–1199.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
