

A novel ring-like complex of Xenopus proteins essential for the initiation of DNA replication
- PMID: 12730133
- PMCID: PMC196047
- DOI: 10.1101/gad.1070003
A novel ring-like complex of Xenopus proteins essential for the initiation of DNA replication
Abstract
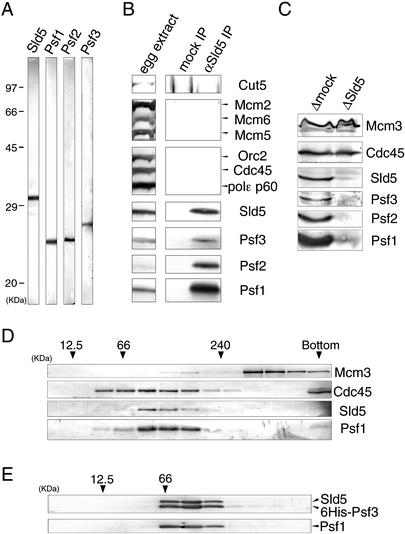
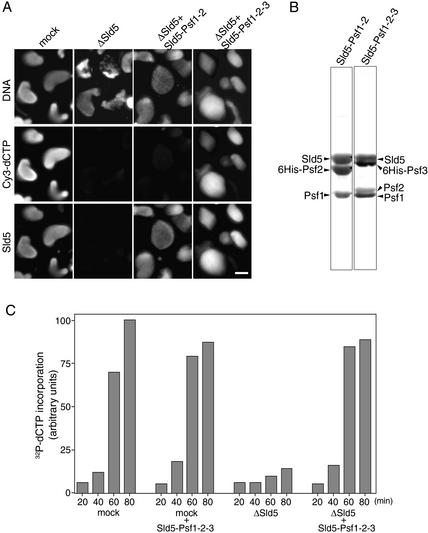
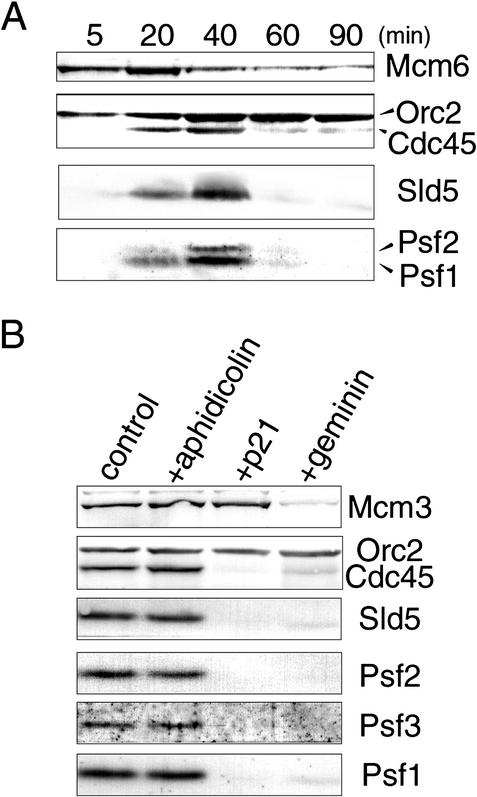
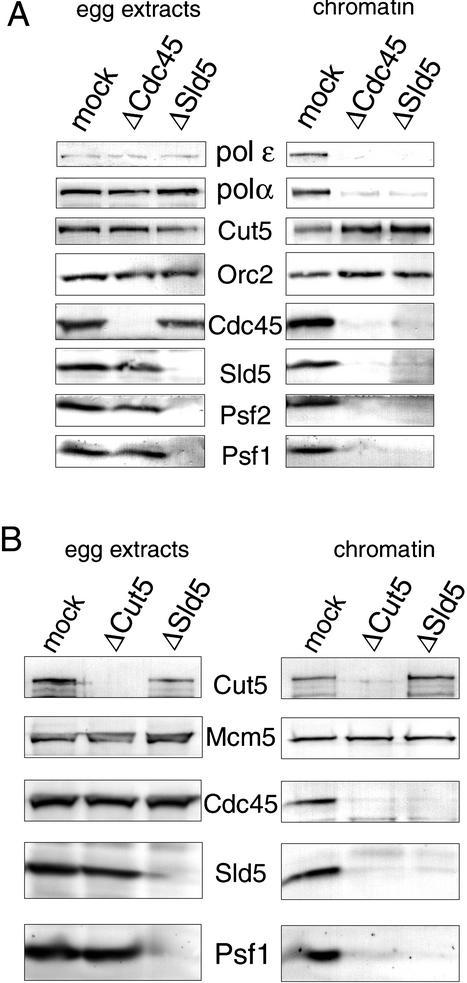
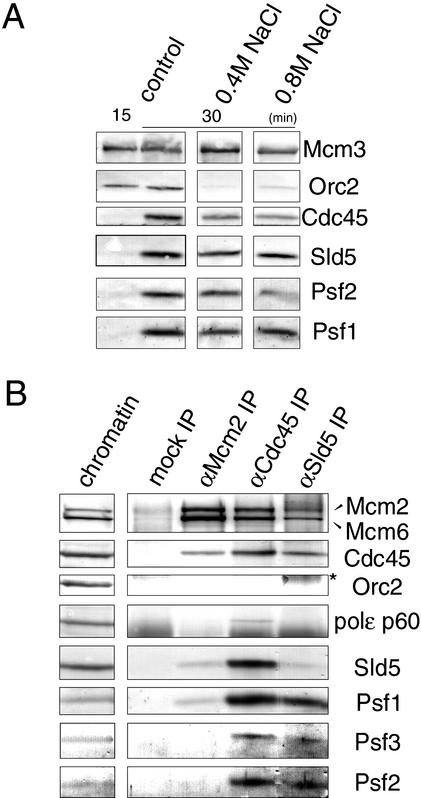
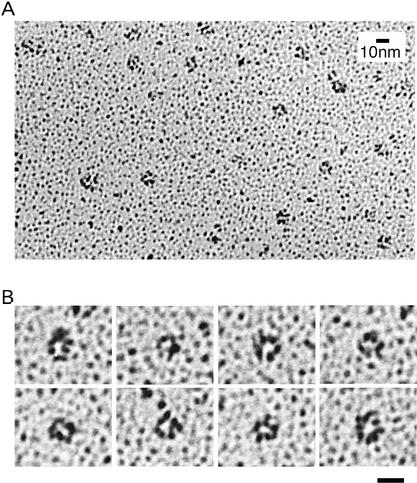
We have identified Xenopus homologs of the budding yeast Sld5 and its three interacting proteins. These form a novel complex essential for the initiation of DNA replication in Xenopus egg extracts. The complex binds to chromatin in a manner dependent on replication licensing and S-phase CDK. The chromatin binding of the complex and that of Cdc45 are mutually dependent and both bindings require Xenopus Cut5, the yeast homolog of which interacts with Sld5. On replicating chromatin the complex interacts with Cdc45 and MCM, putative components of replication machinery. Electron microscopy further reveals that the complex has a ring-like structure. These results suggest that the complex plays an essential role in the elongation stage of DNA replication as well as the initiation stage.
Figures

References
-
- Aparicio OM, Weinstein DM, Bell SP. Components and dynamics of DNA replication complexes in S. cerevisiae: Redistribution of MCM proteins and Cdc45p during S phase. Cell. 1997;91:59–69. - PubMed
-
- Arata T. Electron microscopic observation of monomeric actin attached to a myosin head. J Struct Biol. 1998;123:8–16. - PubMed
-
- Bell SP, Dutta A. DNA replication in eukaryotic cells. Annu Rev Biochem. 2002;71:333–374. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases