

Molecular analysis of the LATERAL SUPPRESSOR gene in Arabidopsis reveals a conserved control mechanism for axillary meristem formation
- PMID: 12730136
- PMCID: PMC196050
- DOI: 10.1101/gad.260703
Molecular analysis of the LATERAL SUPPRESSOR gene in Arabidopsis reveals a conserved control mechanism for axillary meristem formation
Abstract
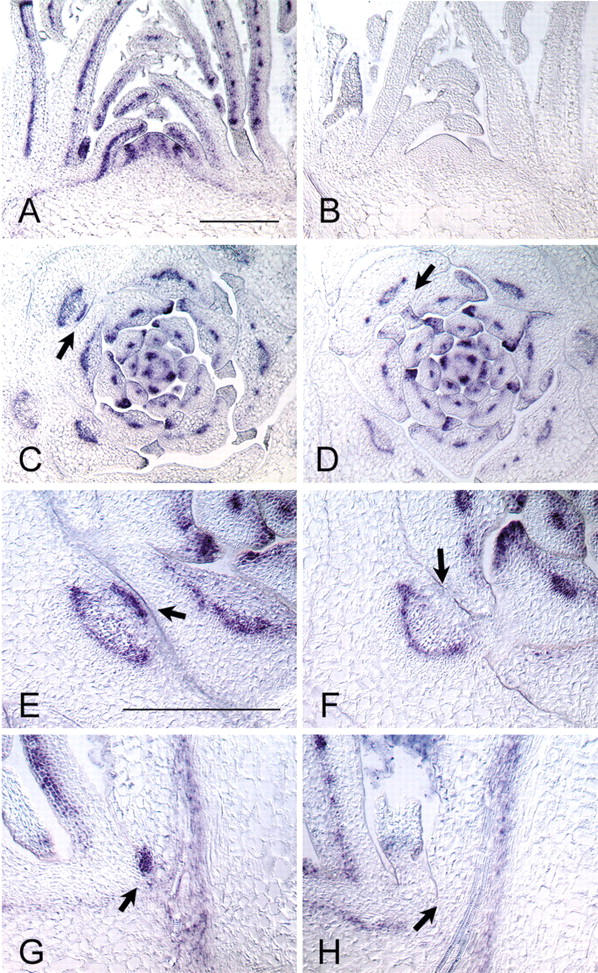
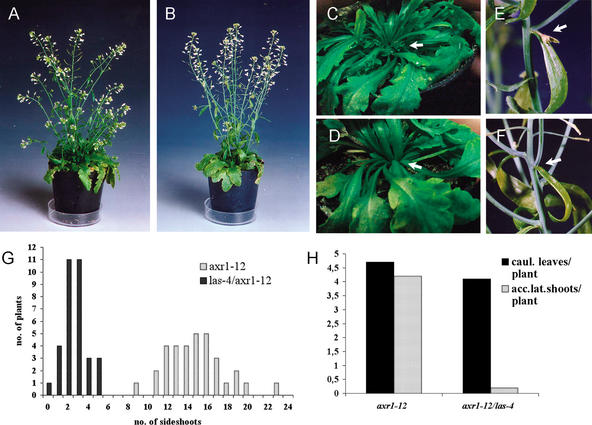
In seed plants, shoot branching is initiated by the formation of new meristems in the axils of leaves, which subsequently develop into new axes of growth. This study describes the genetic control of axillary meristem formation by the LATERAL SUPPRESSOR (LAS) gene in Arabidopsis thaliana. las mutants show a novel phenotype that is characterized by the inability to form lateral shoots during vegetative development. The analysis shows that axillary meristem formation is differently regulated during different phases of development. During reproductive development, axillary meristems initiate in close proximity to the shoot apical meristem and do not require LAS function. In contrast, during the vegetative phase, axillary meristems initiate at a distance to the SAM and require LAS function. This control mechanism is conserved between the distantly related species tomato and Arabidopsis. Monitoring the patterns of LAS and SHOOT MERISTEMLESS transcript accumulation allowed us to identify early steps in the development of leaf axil identity, which seem to be a prerequisite for axillary meristem initiation. Other regulators of shoot branching, like REVOLUTA and AUXIN RESISTANT 1, act downstream of LAS. The results are discussed in the context of the "detached meristem" and the "de novo formation" concepts of axillary meristem formation.
Figures
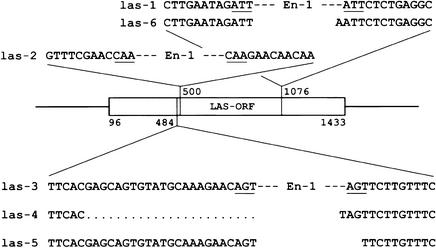


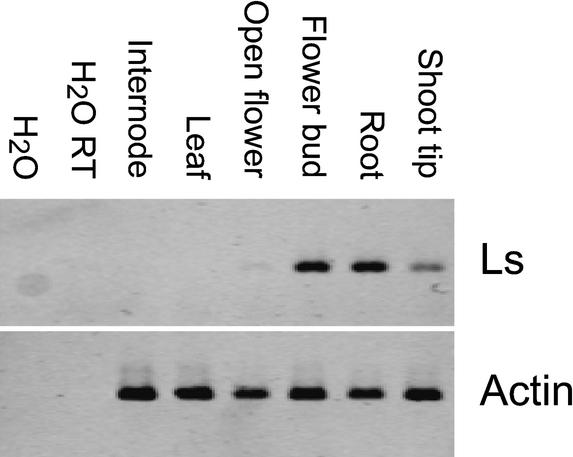
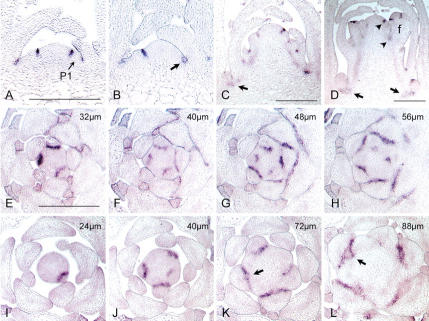
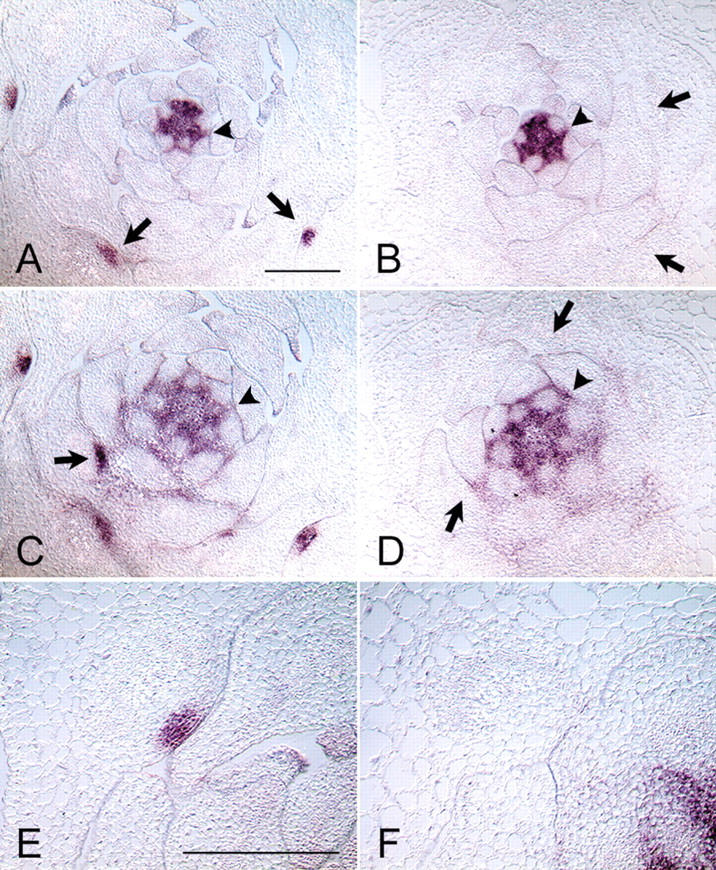
References
-
- Abel S, Nguyen MD, Theologis A. The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana. J Mol Biol. 1995;251:533–549. - PubMed
-
- Aida M, Ishida T, Tasaka M. Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: Interaction among the CUP-SHAPED COTYLEDON and SHOOT MERISTEMLESS genes. Development. 1999;126:1563–1570. - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Baumann E, Lewald J, Saedler H, Schulz B, Wisman E. Successful PCR-based reverse genetic screen using an En-1-mutagenised Arabidopsis thaliana population generated via single-seed descent. Theor Appl Genet. 1998;97:729–734.
-
- Becker D, Kemper E, Schell J, Masterson R. New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Mol Biol. 1992;20:1195–1197. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases