

Identification of rocA, a positive regulator of covR expression in the group A streptococcus
- PMID: 12730168
- PMCID: PMC154078
- DOI: 10.1128/JB.185.10.3081-3090.2003
Identification of rocA, a positive regulator of covR expression in the group A streptococcus
Abstract
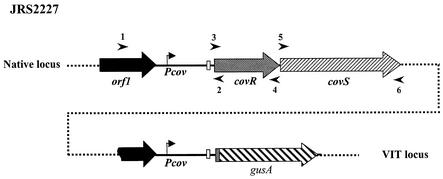
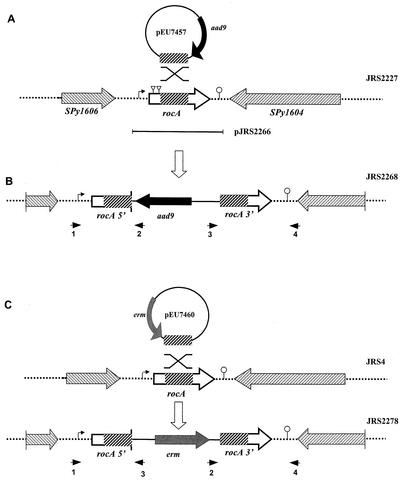
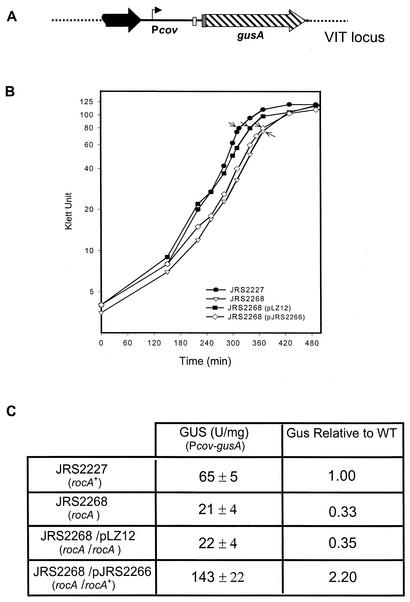
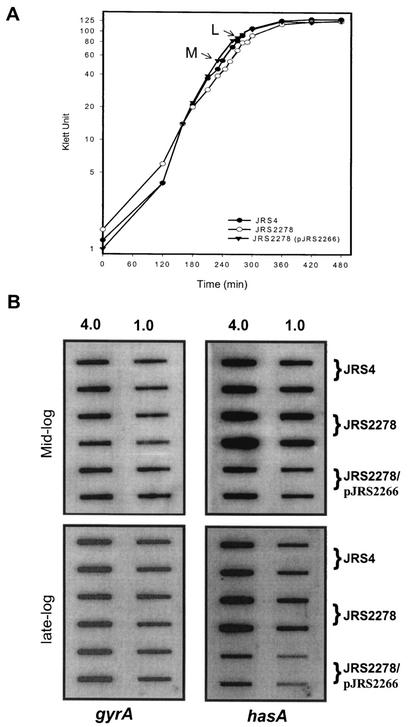
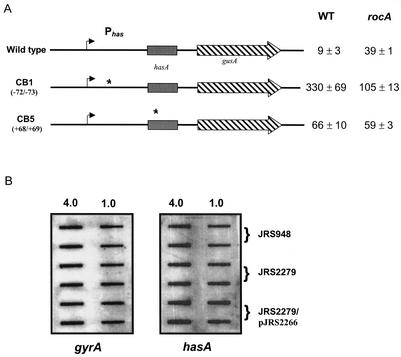
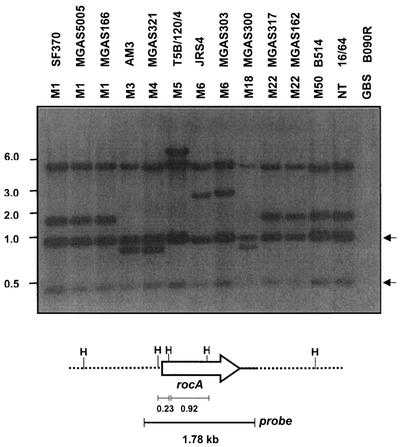
In the group A streptococcus (GAS; Streptococcus pyogenes), a two-component system known as CovRS (or CsrRS) regulates about 15% of the genes, including several important virulence factors like the hyaluronic acid capsule. Most of these genes, including covR itself, are negatively regulated by CovR. We have isolated two independent ISS1 insertions in an open reading frame (ORF) that increases CovR expression as measured by a Pcov-gusA reporter fusion in single copy in the GAS chromosome. This ORF, named rocA for "regulator of Cov," activates covR transcription about threefold. As expected, a rocA mutant is mucoid and produces more transcript from the has promoter since this promoter is repressed by CovR. This effect is dependent on the presence of a wild-type covR gene. In contrast to its activation of Pcov, RocA negatively regulates its own expression. This autoregulation is not dependent on the presence of the covR gene. All the phenotypes of the rocA mutant were complemented by the presence of the rocA gene on a plasmid. The rocA gene is present in strains of all nine M serotypes of GAS tested and is absent from strains representing 11 other groups of streptococci and related bacteria, including strains of the closely related group C and G streptococci. It seems likely that rocA plays an important role in the pathogenesis of GAS since it affects expression of the global regulator CovR.
Figures

References
-
- Alouf, J., and H. Muller-Alouf. 1996. Cellular constituents and extracellular proteins involved in the pathogenic capacity of Streptococcus group A. Ann. Pharm. Fr. 54:49-59. - PubMed
-
- Bernish, B., and I. van de Rijn. 1999. Characterization of a two-component system in Streptococcus pyogenes which is involved in regulation of hyaluronic acid production. J. Biol. Chem. 274:4786-4793. - PubMed
-
- Bisno, A. L. 1991. Group A streptococcal infections and acute rheumatic fever. N. Engl. J. Med. 325:783-793. - PubMed
-
- Bronze, M. S., and J. B. Dale. 1996. The reemergence of serious group A streptococcal infections and acute rheumatic fever. Am. J. Med. Sci. 311:41-54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
