

Substoichiometric shifting in the plant mitochondrial genome is influenced by a gene homologous to MutS
- PMID: 12730382
- PMCID: PMC156310
- DOI: 10.1073/pnas.1037651100
Substoichiometric shifting in the plant mitochondrial genome is influenced by a gene homologous to MutS
Abstract
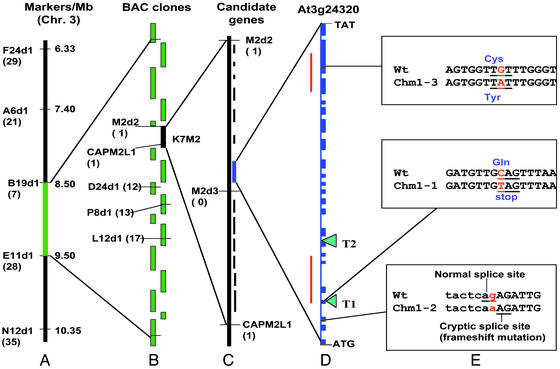
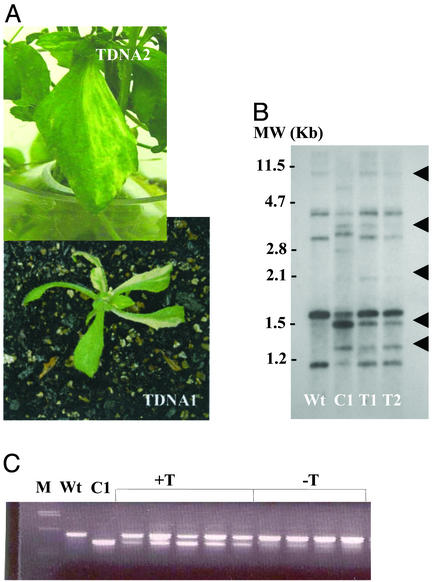
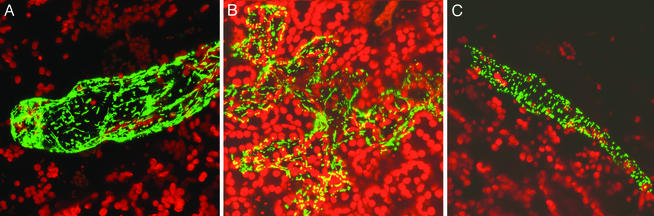
The plant mitochondrial genome is retained in a multipartite structure that arises by a process of repeat-mediated homologous recombination. Low-frequency ectopic recombination also occurs, often producing sequence chimeras, aberrant ORFs, and novel subgenomic DNA molecules. This genomic plasticity may distinguish the plant mitochondrion from mammalian and fungal types. In plants, relative copy number of recombination-derived subgenomic DNA molecules within mitochondria is controlled by nuclear genes, and a genomic shifting process can result in their differential copy number suppression to nearly undetectable levels. We have cloned a nuclear gene that regulates mitochondrial substoichiometric shifting in Arabidopsis. The CHM gene was shown to encode a protein related to the MutS protein of Escherichia coli that is involved in mismatch repair and DNA recombination. We postulate that the process of substoichiometric shifting in plants may be a consequence of ectopic recombination suppression or replication stalling at ectopic recombination sites to effect molecule-specific copy number modulation.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
