

Bandpass filtering at the rod to second-order cell synapse in salamander (Ambystoma tigrinum) retina
- PMID: 12736350
- PMCID: PMC6742206
- DOI: 10.1523/JNEUROSCI.23-09-03796.2003
Bandpass filtering at the rod to second-order cell synapse in salamander (Ambystoma tigrinum) retina
Abstract
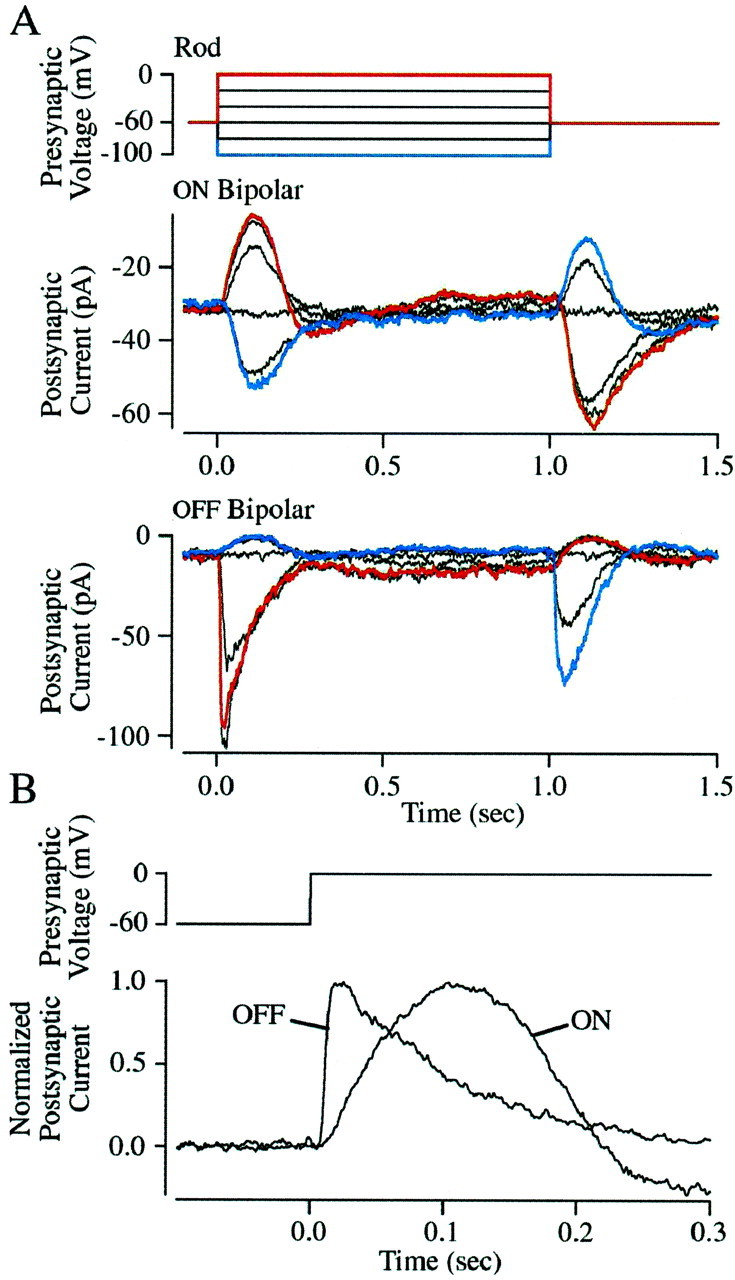
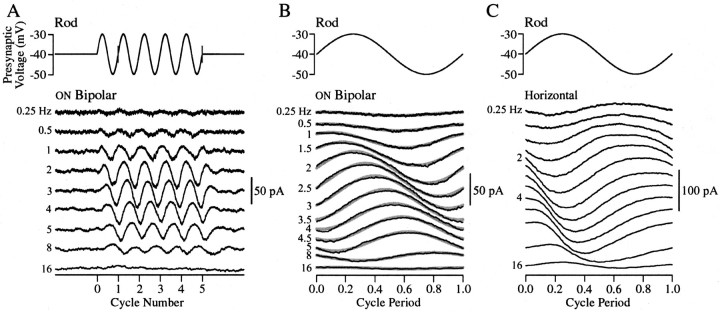
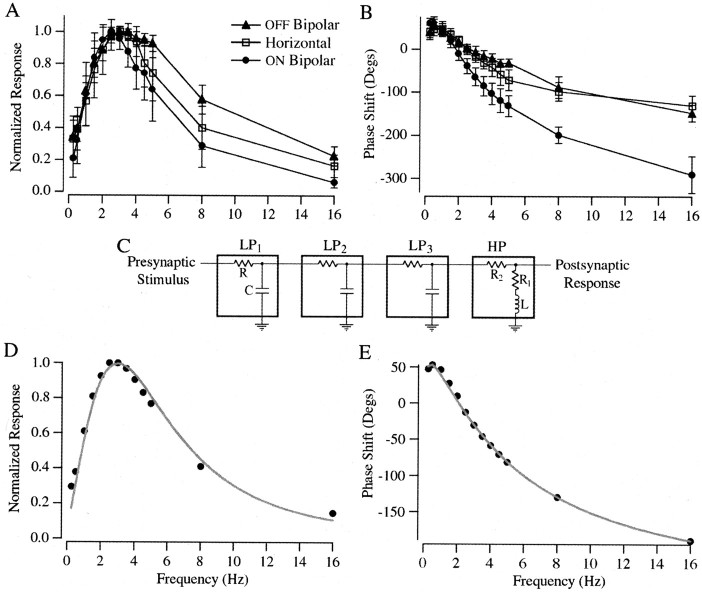
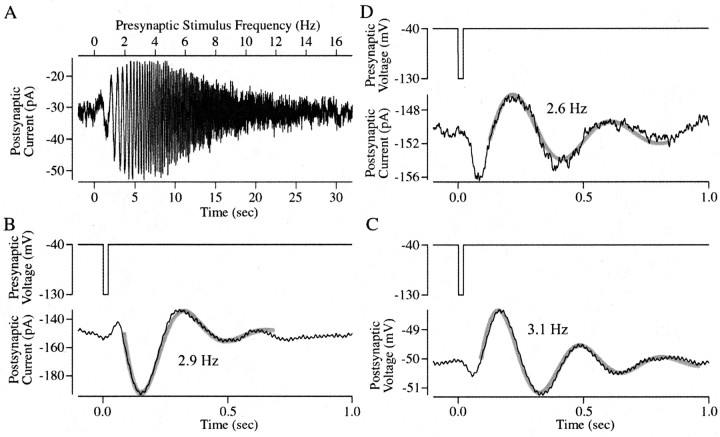
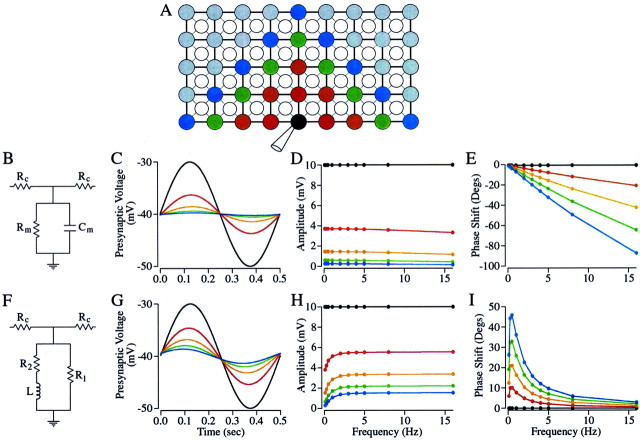
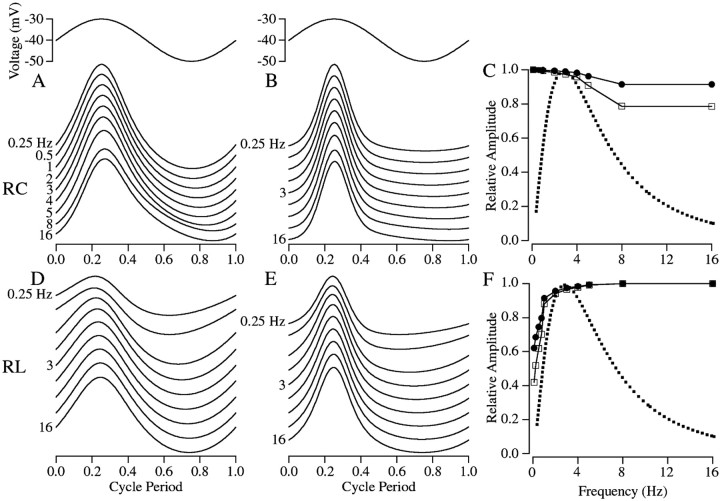
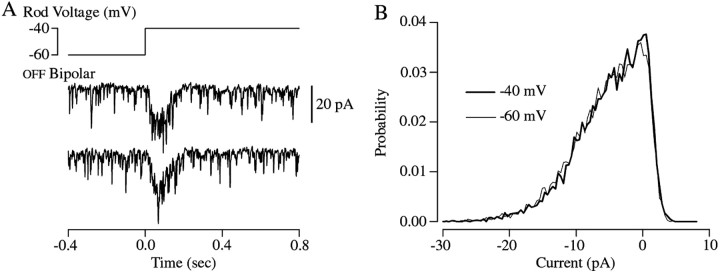
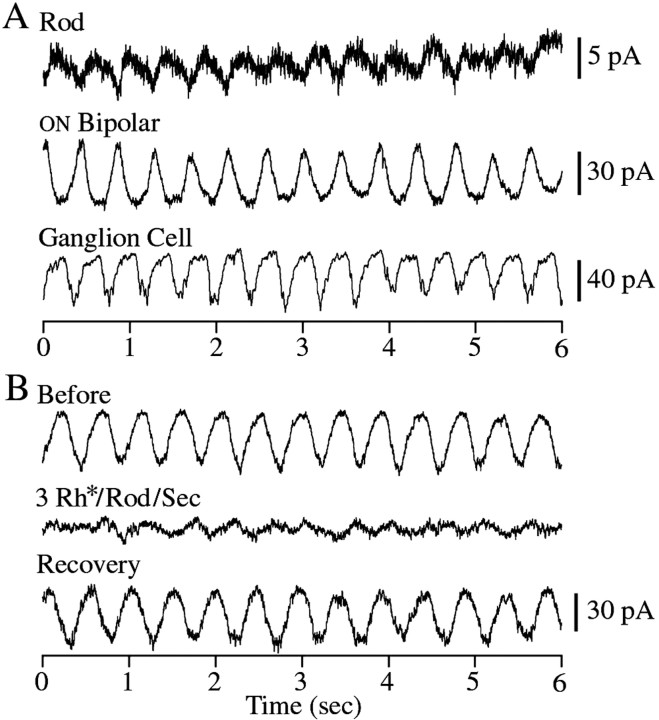
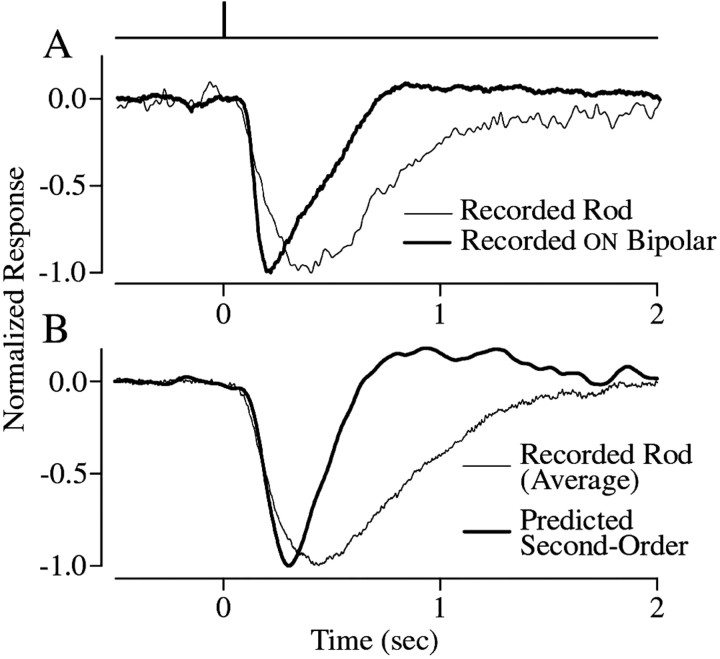
The ability to see at night relies on the transduction of single photons by the rod photoreceptors and transmission of the resulting signals through the retina. Using paired patch-clamp recordings, we investigated the properties of the first stage of neural processing of the rod light responses: signal transfer from rods to bipolar and horizontal cells. Bypassing the relatively slow phototransduction process and directly modulating the rod voltage or current allowed us to characterize signal transfer over a wide range of temporal frequencies. We found that the rod to second-order cell synapse acts as a bandpass filter, preferentially transmitting signals with frequencies between 1.5 and 4 Hz while attenuating higher and lower frequency inputs. The similarity of the responses in different types of postsynaptic cell and the properties of miniature EPSCs (mEPSCs) recorded in OFF bipolar cells suggest that most of the bandpass filtering is mediated presynaptically. Modeling of the network of electrically coupled rod photoreceptors suggests that spread of the signal through the network contributed to the observed high-pass filtering but not to the low-pass filtering. Attenuation of low temporal frequencies at the first retinal synapse sharpens the temporal resolution of the light response; attenuation of high temporal frequencies removes voltage noise in the rod that threatens to swamp the light response.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources