

Neural correlates of competing fear behaviors evoked by an innately aversive stimulus
- PMID: 12736356
- PMCID: PMC6742203
- DOI: 10.1523/JNEUROSCI.23-09-03855.2003
Neural correlates of competing fear behaviors evoked by an innately aversive stimulus
Abstract
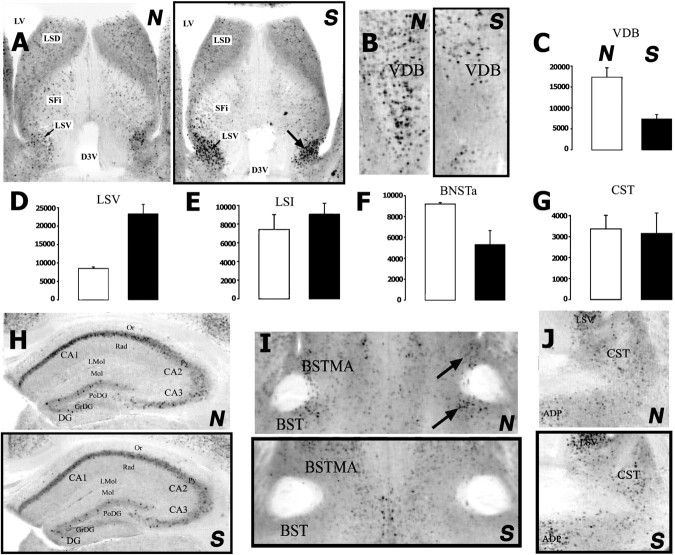
Environment and experience influence defensive behaviors, but the neural circuits mediating such effects are not well understood. We describe a new experimental model in which either flight or freezing reactions can be elicited from mice by innately aversive ultrasound. Flight and freezing are negatively correlated, suggesting a competition between fear motor systems. An unfamiliar environment or a previous aversive event, moreover, can alter the balance between these behaviors. To identify potential circuits controlling this competition, global activity patterns in the whole brain were surveyed in an unbiased manner by c-fos in situ hybridization, using novel experimental and analytical methods. Mice predominantly displaying freezing behavior had preferential neural activity in the lateral septum ventral and several medial and periventricular hypothalamic nuclei, whereas mice predominantly displaying flight had more activity in cortical, amygdalar, and striatal motor areas, the dorsolateral posterior zone of the hypothalamus, and the vertical limb of the diagonal band. These complementary patterns of c-fos induction, taken together with known connections between these structures, suggest ways in which the brain may mediate the balance between these opponent defensive behaviors.
Figures







References
-
- Alheid GF, Olmos JS, Beltramino CA. Amygdala and extended amygdala. In: Paxinos G, editor. The rat nervous system. Academic; San Diego: 1995. pp. 495–578.
-
- Anokhin KV, Mileusnic R, Shamakina IY, Rose SP. Effects of early experience on c-fos gene expression in the chick forebrain. Brain Res. 1991;544:101–107. - PubMed
-
- Bandler R, Keay KA, Floyd N, Price J. Central circuits mediating patterned autonomic activity during active vs. passive emotional coping. Brain Res Bull. 2000;53:95–104. - PubMed
-
- Beckett SR, Aspley S, Graham M, Marsden CA. Pharmacological manipulation of ultrasound induced defence behaviour in the rat. Psychopharmacology. 1996;127:384–390. - PubMed
-
- Blanchard DC, Griebel G, Blanchard RJ. Mouse defensive behaviors: pharmacological and behavioral assays for anxiety and panic. Neurosci Biobehav Rev. 2001;25:205–218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources