

Fatty acid synthesis is essential in embryonic development: fatty acid synthase null mutants and most of the heterozygotes die in utero
- PMID: 12738878
- PMCID: PMC164451
- DOI: 10.1073/pnas.0931394100
Fatty acid synthesis is essential in embryonic development: fatty acid synthase null mutants and most of the heterozygotes die in utero
Abstract
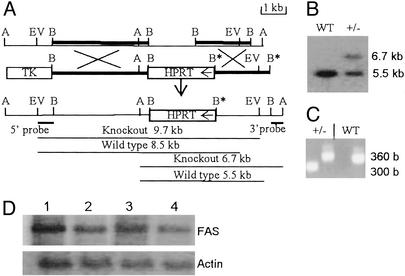
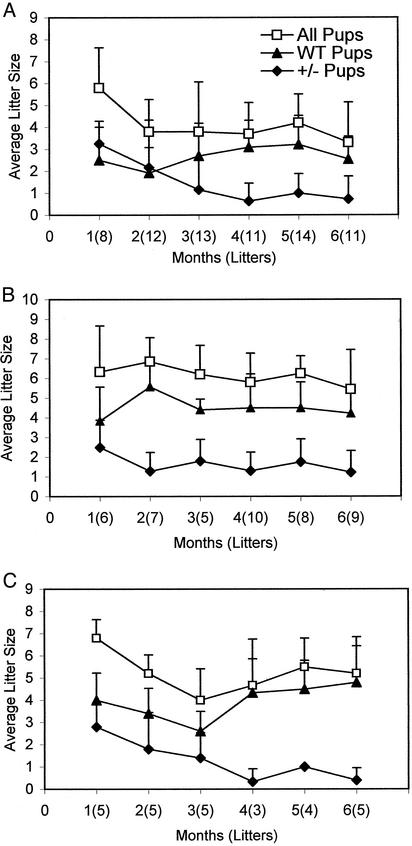
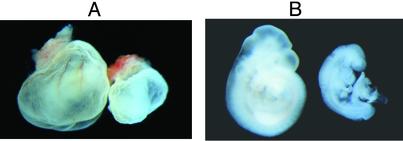
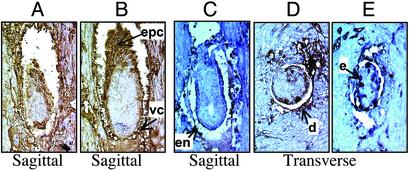
In animals, including humans, the source of long-chain saturated fatty acids is de novo synthesis, which is mediated by fatty acid synthase (FAS), ingested food, or both. To understand the importance of de novo fatty acid synthesis, we generated FAS knockout mice. The heterozygous FAS mutants (Fasn+/-) are ostensibly normal. In Fasn+/- mice the levels of FAS mRNA and the FAS activity are approximately 50% and 35% lower, respectively, than those of WT mice; hence, FAS levels are affected by gene dosage. When the Fasn+/- mutant mice were interbred, Fasn-/- mice were not produced; thus, FAS is essential during embryonic development. Furthermore, the number of Fasn+/- progeny obtained was 70% less than predicted by Mendelian inheritance, indicating partial haploid insufficiency. Even when one of the parents was WT, the estimated loss of heterozygous progeny was 60%. This loss of Fasn+/- pups appeared to be strain-specific and became more pronounced as the heterozygous females produced more litters. Most of the Fasn-/- mutant embryos died before implantation and the Fasn+/- embryos died at various stages of their development. Feeding the breeders a diet rich in saturated fatty acids did not prevent the loss of homoor heterozygotes. These observations are very important in considering teratogenic consequences of drugs aimed at inhibiting FAS activity, to reduce either obesity or the growth of cancerous tissues.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
