

Space-time maps and two-bar interactions of different classes of direction-selective cells in macaque V-1
- PMID: 12740411
- PMCID: PMC2627780
- DOI: 10.1152/jn.00550.2002
Space-time maps and two-bar interactions of different classes of direction-selective cells in macaque V-1
Abstract
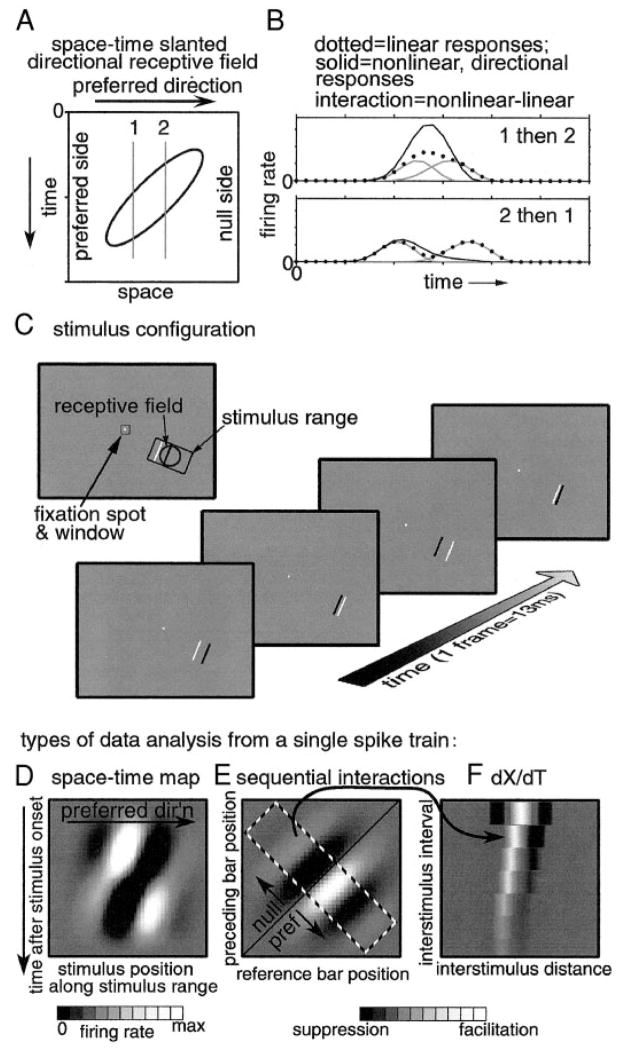
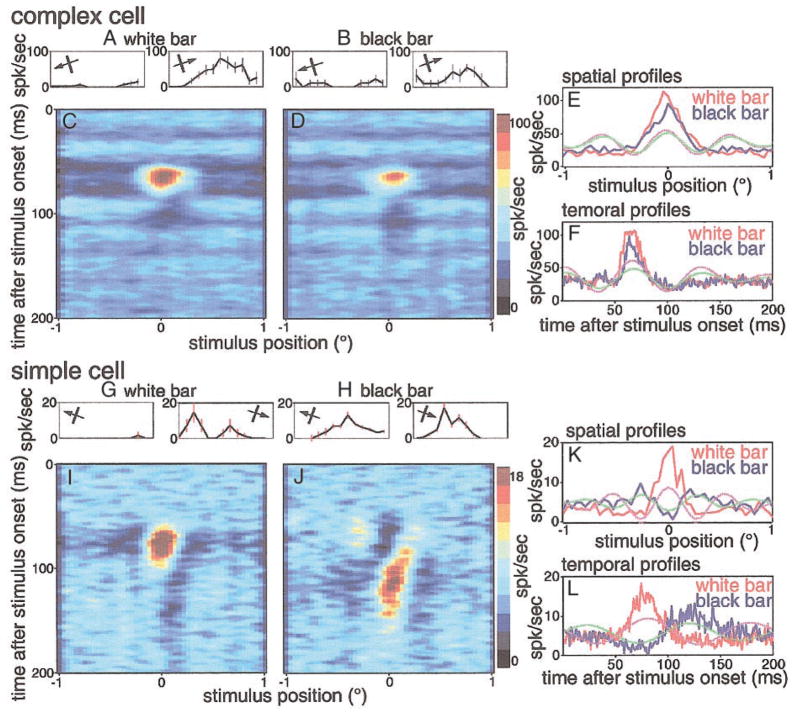
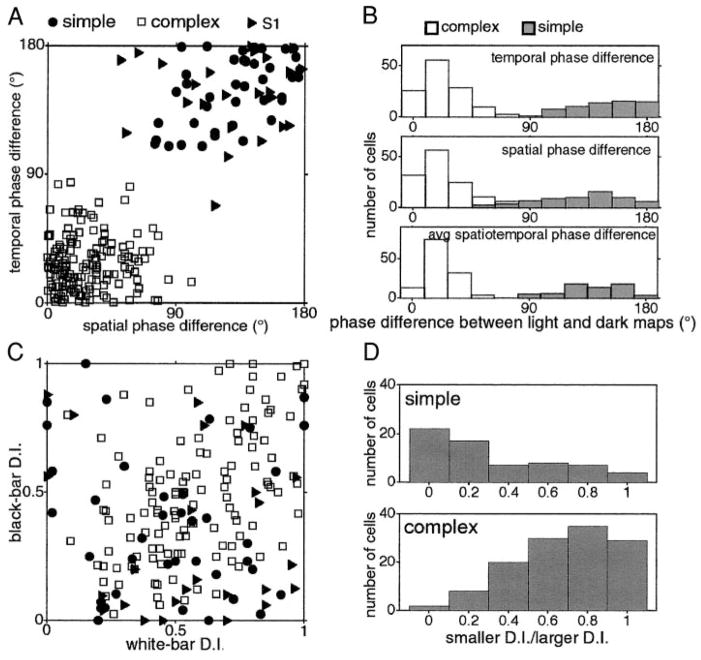
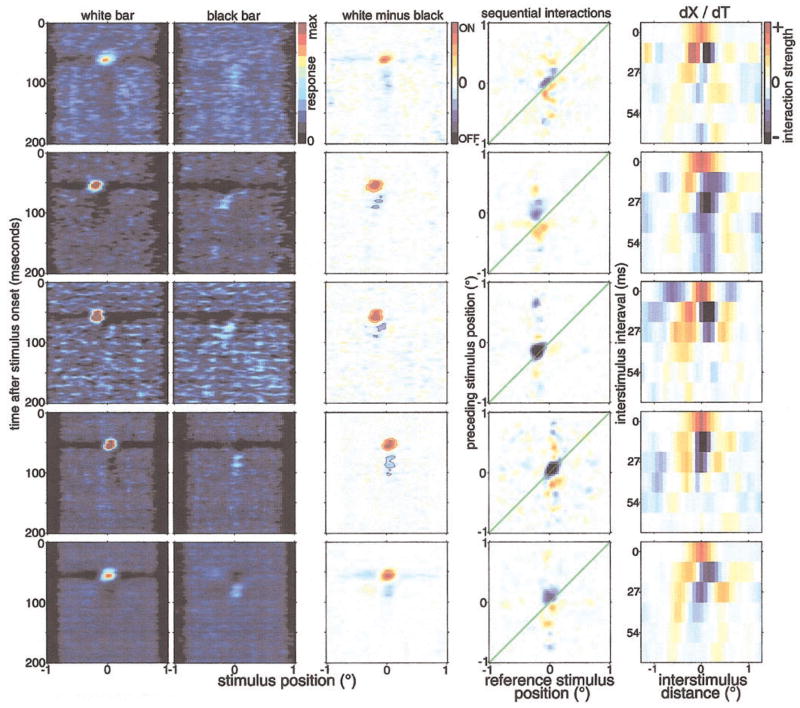
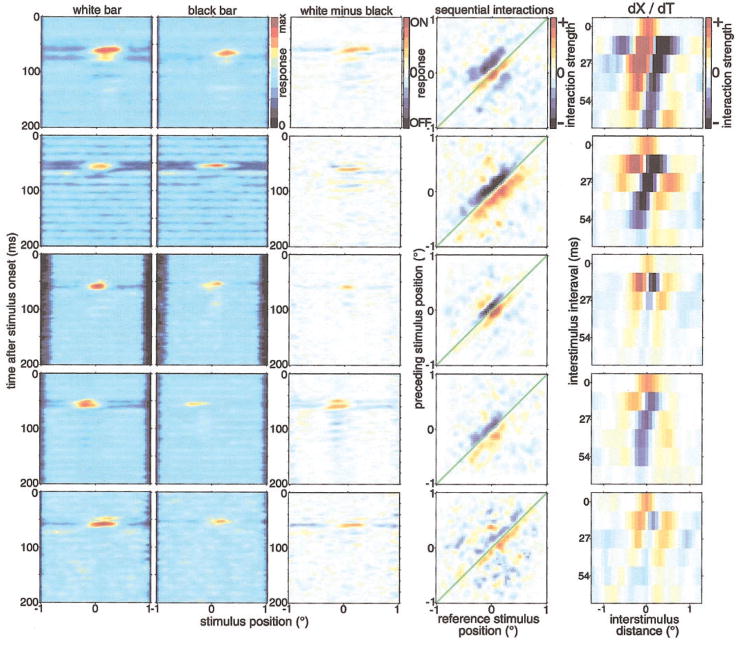
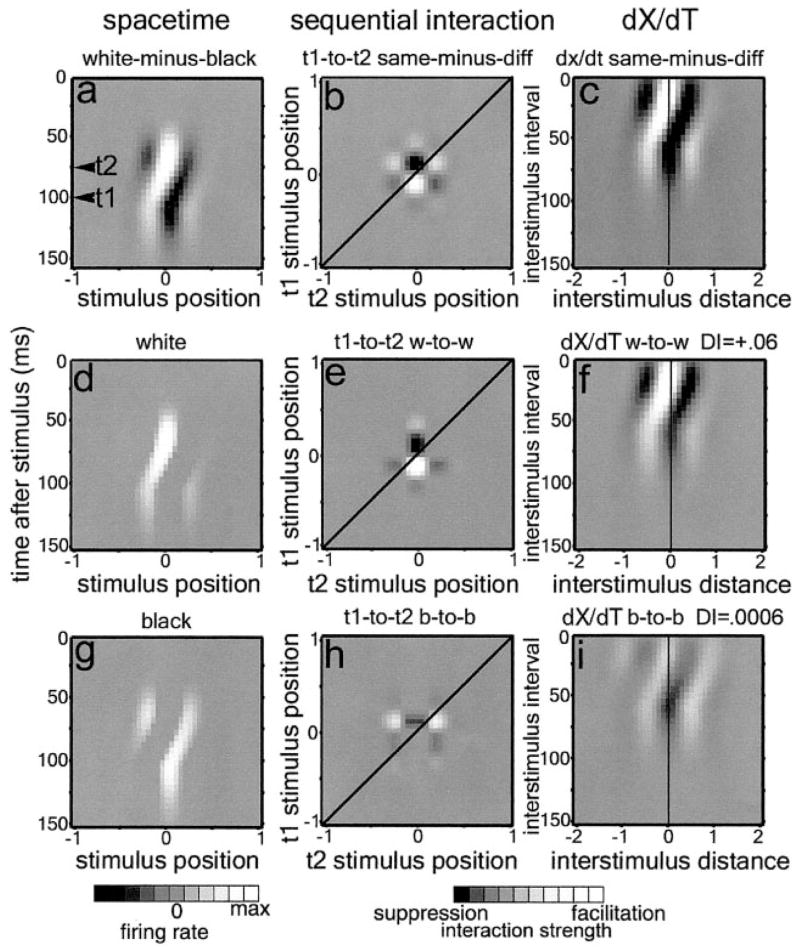
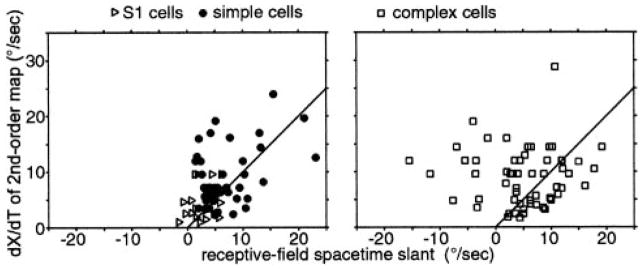
We used one-dimensional sparse noise stimuli to generate first-order spatiotemporal maps and second-order two-bar interaction maps for 65 simple and 124 complex direction-selective cells in alert macaque V1. Spatial and temporal phase differences between light and dark space-time maps clearly distinguished simple and complex cell populations. Complex cells usually showed similar direction preferences to light and dark bars, but many of the directional simple cells were much more direction selective to one sign of contrast than the reverse. We show that this is predicted by a simple energy model. Some of the direction-selective simple cells showed multiple space-time-slanted subregions, but others (previously described as S1 cells) had space-time maps that looked like just one subregion of an ordinary simple cell. Both simple and complex cells showed directional interactions (nonlinearities) to pairs of flashed bars (a 2-bar apparent-motion stimulus). The space-time slant of the simple cells correlated with the optimum dX/dT (velocity) of the paired-bar interactions. Some complex cells also showed a space-time slant; the direction of the slant usually correlated with the preferred direction of motion, but the degree of slant correlated with the inferred velocity tuning only when measured by a weighted-centroid calculation. Principal components analysis of the simple-cell space-time maps yielded one fast temporally biphasic component and a slower temporally monophasic component. We saw no consistent pattern for the spatial phase of the components, unlike previous reports; however, we show that principal components analysis may not distinguish between spatial offsets and phase offsets.
Figures





Similar articles
-
Substructure of direction-selective receptive fields in macaque V1.J Neurophysiol. 2003 May;89(5):2743-59. doi: 10.1152/jn.00822.2002. J Neurophysiol. 2003. PMID: 12740412 Free PMC article.
-
Contrast affects speed tuning, space-time slant, and receptive-field organization of simple cells in macaque V1.J Neurophysiol. 2007 Jan;97(1):849-57. doi: 10.1152/jn.00762.2006. Epub 2006 Nov 15. J Neurophysiol. 2007. PMID: 17108092 Free PMC article.
-
Direction selectivity of simple cells in cat striate cortex to moving light bars. II. Relation to moving dark bar responses.Exp Brain Res. 1985;57(3):523-36. doi: 10.1007/BF00237839. Exp Brain Res. 1985. PMID: 3979495
-
Visual properties of neurons in area V4 of the macaque: sensitivity to stimulus form.J Neurophysiol. 1987 Mar;57(3):835-68. doi: 10.1152/jn.1987.57.3.835. J Neurophysiol. 1987. PMID: 3559704
-
Directional inhibition: a new slant on an old question.Neuron. 2005 Jan 6;45(1):5-7. doi: 10.1016/j.neuron.2004.12.029. Neuron. 2005. PMID: 15629696 Review.
Cited by
-
Perceptual consequences of feature-based attentional enhancement and suppression.J Vis. 2012 Aug 24;12(8):15. doi: 10.1167/12.8.15. J Vis. 2012. PMID: 22923726 Free PMC article.
-
The profile of attentional modulation to visual features.J Vis. 2019 Nov 1;19(13):13. doi: 10.1167/19.13.13. J Vis. 2019. PMID: 31747691 Free PMC article.
-
Synaptic mechanisms of direction selectivity in primary auditory cortex.J Neurosci. 2010 Feb 3;30(5):1861-8. doi: 10.1523/JNEUROSCI.3088-09.2010. J Neurosci. 2010. PMID: 20130195 Free PMC article.
-
Mapping receptive fields in primary visual cortex.J Physiol. 2004 Aug 1;558(Pt 3):717-28. doi: 10.1113/jphysiol.2004.065771. Epub 2004 May 21. J Physiol. 2004. PMID: 15155794 Free PMC article. Review.
-
Responses of V1 neurons to two-dimensional hermite functions.J Neurophysiol. 2006 Jan;95(1):379-400. doi: 10.1152/jn.00498.2005. Epub 2005 Sep 7. J Neurophysiol. 2006. PMID: 16148274 Free PMC article.
References
-
- Adelson EH, Bergen JR. Spatiotemporal energy models for the perception of motion. J Opt Soc Am A Opt Image Sci. 1985;2:284–299. - PubMed
-
- Albrecht DG, Geisler WS. Motion selectivity and the contrast-response function of simple cells in the visual cortex. Vis Neurosc. 1991;7:531–546. - PubMed
-
- Baker CL., Jr Linear filtering and nonlinear interactions in direction-selective visual cortex neurons: a noise correlation analysis. Vis Neurosci. 2001;18:465–485. - PubMed
-
- Baker CL, Cynader MS. Space-time separability of direction selectivity in cat striate cortex neurons. Vision Res. 1988;28:239–246. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources